Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
N |
|---|
náboj amfiontuangl. amphion charge, součet nábojů jednotlivých ionisovaných skupin v 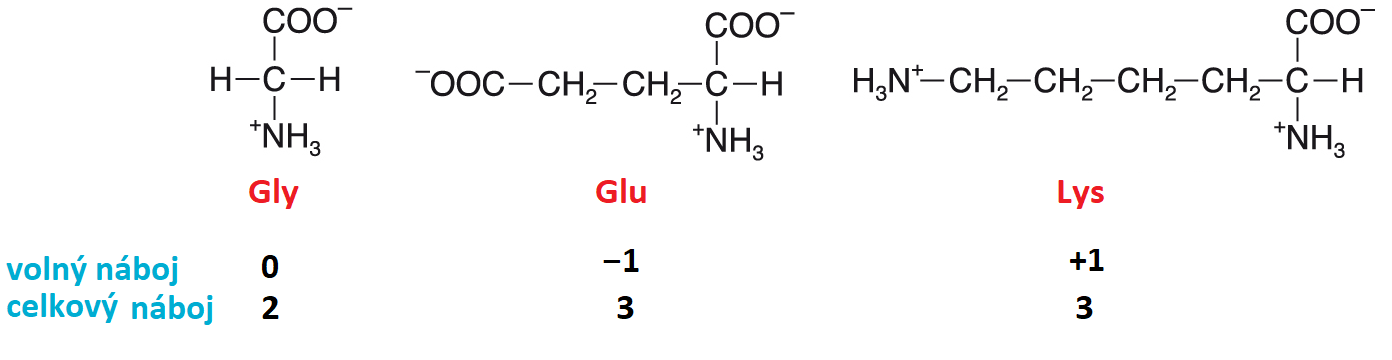 molekule s přihlédnutím k jejich znaménku (též volný náboj); někdy se udává i celkový náboj, což je počet nábojů amfiontu bez ohledu na jejich znaménko. Na obrázku jsou zachyceny náboje pro neutrální, kyselou a bazickou aminokyselinu ve vodném roztoku při pH 7. molekule s přihlédnutím k jejich znaménku (též volný náboj); někdy se udává i celkový náboj, což je počet nábojů amfiontu bez ohledu na jejich znaménko. Na obrázku jsou zachyceny náboje pro neutrální, kyselou a bazickou aminokyselinu ve vodném roztoku při pH 7. | |
náboj buňky energetickýangl. cell energy charge, veličina vyjadřující stav energetických rezerv buňky:
energetický náboj buňky\( = \frac{[ATP]+ \frac{1}{2} [ADP]}{[ATP]+[ADP]+[AMP]} \) Faktor ½ pro koncentraci ADP ukazuje, že jeho štěpení na AMP poskytuje polovinu energie odpovídající štěpení ATP (viz adenosinfosfáty, energetická jednotka ATP). Energetický náboj může teoreticky dosáhnout hodnot mezi 0 (adenosinfosfáty jsou přítomny pouze jako AMP) až 1 (jsou přítomny pouze jako ATP). | |
NADnikotinamidadenindinukleotid, angl. nicotinamide adenine 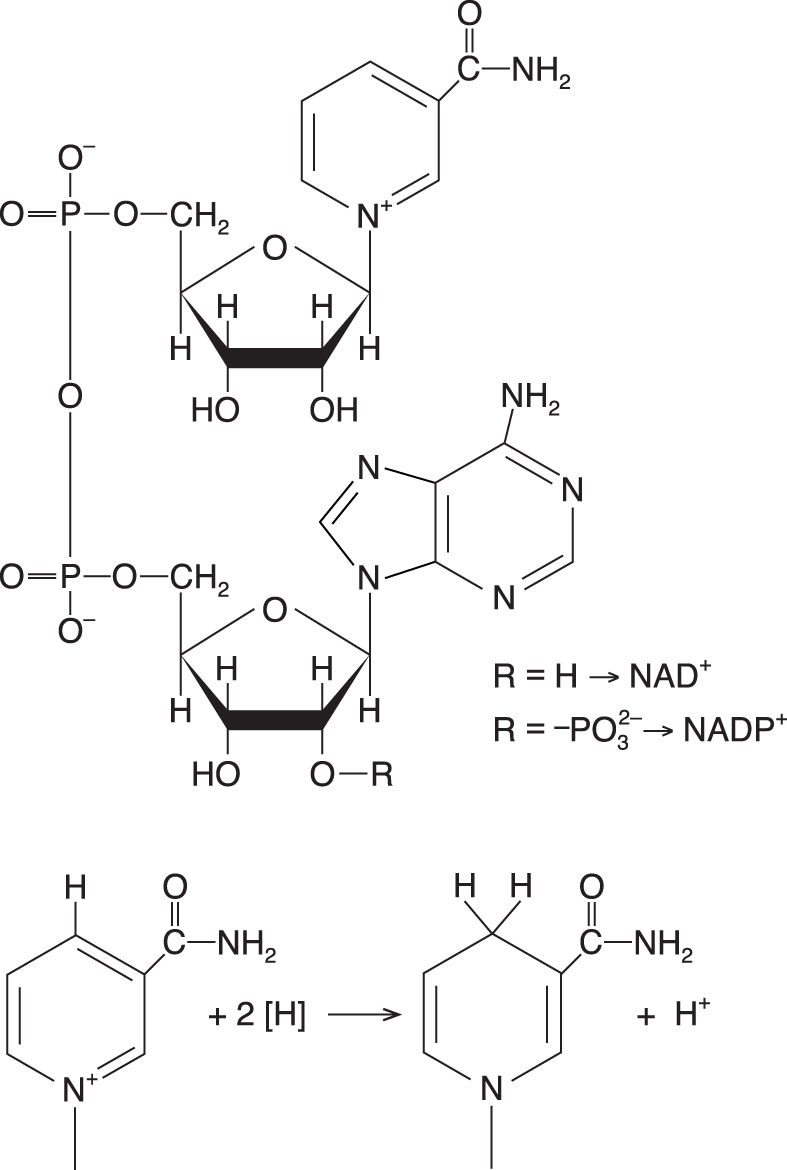 dinucleotide, starší označení difosfopyridin-nukleotid (zkratka DPN), jeden z nejvýznamnějších
koenzymů oxidoreduktas. Protože jeho pyridinové jádro je kladně nabité, označuje se přesněji NAD+; redukcí může přijmout dva elektrony a jeden vodíkový ion (H+) a přejít na formu NADH. Jeho součástí je amid nikotinové kyseliny, vitamin B3 – niacin. Jeho hlavním úkolem v metabolismu je přijímat vodíkové atomy (resp. elektrony) v dehydrogenačních katabolických reakcích (redukce NAD+ na NADH) a přenášet je do dýchacího řetězce, kde se NAD+ regeneruje. Podobné chemické vlastnosti má NADP. Více viz redoxní systém NAD(P)+, NAD(P)H. dinucleotide, starší označení difosfopyridin-nukleotid (zkratka DPN), jeden z nejvýznamnějších
koenzymů oxidoreduktas. Protože jeho pyridinové jádro je kladně nabité, označuje se přesněji NAD+; redukcí může přijmout dva elektrony a jeden vodíkový ion (H+) a přejít na formu NADH. Jeho součástí je amid nikotinové kyseliny, vitamin B3 – niacin. Jeho hlavním úkolem v metabolismu je přijímat vodíkové atomy (resp. elektrony) v dehydrogenačních katabolických reakcích (redukce NAD+ na NADH) a přenášet je do dýchacího řetězce, kde se NAD+ regeneruje. Podobné chemické vlastnosti má NADP. Více viz redoxní systém NAD(P)+, NAD(P)H. | |
NADPnikotinamidadenindinukleotidfosfát, angl. nicotinamide adenine dinucleotide phosphate, fosforylovaný analog
NAD (vzorec viz tamtéž). V redukované formě (NADPH) ho buňky používají jako redukční činidlo v biosyntézách (v temné fázi fotosyntézy, při biosyntéze mastných kyselin apod.); proto se o NADPH často hovoří jako o redukčním ekvivalentu. Vzniká redukcí NADP+ ve světlé fázi fotosyntézy, v dehydrogenační fázi pentosového cyklu, v pyruvát-malátovém cyklu (působením jablečného enzymu na malát) a v některých dalších dehydrogenačních reakcích. Více viz redoxní systém NAD(P)+, NAD(P)H. | |
Na,K-ATPasatéž
Na+/K+-pumpa, systematický název ATP-fosfohydrolasa 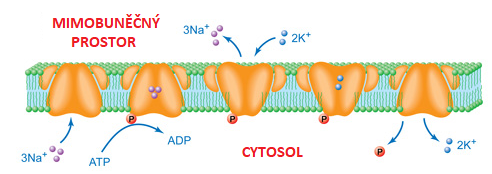 (P-typ, Na+/K+-vyměňující), EC 7.2.2.13, angl. Na,K-ATPase, integrální membránový enzym ze třídy translokas, zajišťující protisměrný primární aktivní transport iontů Na+ a K+. Na úkor hydrolýzy jedné molekuly ATP transportuje tři ionty Na+ z buňky a dva ionty K+ do buňky. Je přítomna ve všech plasmatických membránách, neboť nerovnováha intracelulárních a extracelulárních koncentrací alkalických iontů patří k základním znakům živých buněk; specifické fyziologické role má zejména v buňkách nervových a ledvinových. Za prvotní objev transportního enzymu Na,K–ATPasy obdržel roku 1997 J. C. Skou Nobelovu cenu. (P-typ, Na+/K+-vyměňující), EC 7.2.2.13, angl. Na,K-ATPase, integrální membránový enzym ze třídy translokas, zajišťující protisměrný primární aktivní transport iontů Na+ a K+. Na úkor hydrolýzy jedné molekuly ATP transportuje tři ionty Na+ z buňky a dva ionty K+ do buňky. Je přítomna ve všech plasmatických membránách, neboť nerovnováha intracelulárních a extracelulárních koncentrací alkalických iontů patří k základním znakům živých buněk; specifické fyziologické role má zejména v buňkách nervových a ledvinových. Za prvotní objev transportního enzymu Na,K–ATPasy obdržel roku 1997 J. C. Skou Nobelovu cenu. | |
Nathans Daniel
| |

Neher Erwin
| |

nekrózaangl. necrosis, neprogramovaná (patologická) smrt buňky, ke které dochází v důsledku nevratného narušení některé ze základních buněčných funkcí, např. poškozením biomembrán nebo zástavou oxidační fosforylace, takže dojde k energetickému vyčerpání. Důsledkem nekrózy je bobtnání, následné prasknutí buňky a vylití cytoplasmy do okolí, což může způsobovat zánětlivou reakci (srov. apoptóza). | |
neuromodulátoryangl. neuromodulators, endogenní neuroaktivní
látky, které působí na větší oblast mozku a mnohem pomaleji než neurotransmitery. Nejčastěji ovlivňují nervové buňky prostřednictvím receptorů spřažených s G-proteiny, zatímco neurotransmitery působí prostřednictvím chemicky řízených iontových kanálů. Nejznámější skupinou neuromodulátorů jsou endorfiny, podobně však působí noradrenalin, dopamin, serotonin a acetylcholin. | |
neurotransmiterytéž nervové mediátory, angl. neurotransmitters, nízkomolekulární látky zajišťující jednosměrné předání vzruchu (impulsu) z řídící nervové (presynaptické) buňky do buňky řízené, kterou může být buňka nervová, svalová či žlázová. Jsou vylučovány procesem
exocytózy z presynaptické buňky a difundují synaptickou štěrbinou k membráně postsynaptické buňky, kde se nekovalentně vážou na receptory a vyvolávají buněčnou odpověď (viz synapse). Mezi nejdůležitější neurotransmitery patří acetylcholin, aminokyseliny glycin a glutamová kyselina, katecholaminy dopamin, noradrenalin a adrenalin a biogenní aminy γ-aminomáselná kyselina (GABA), β-alanin, histamin a serotonin. Některé neurotransmitery se užívají jako léčiva při závažných neurologických chorobách (Parkinsonově chorobě) a psychiatrických onemocněních (endogenní depresi, schizofrénii apod.). | |
N-formylmethioninangl. N-formylmethionine, derivát
methioninu, který zahajuje biosyntézu peptidového řetězce u prokaryot. Vzniká postsyntetickou 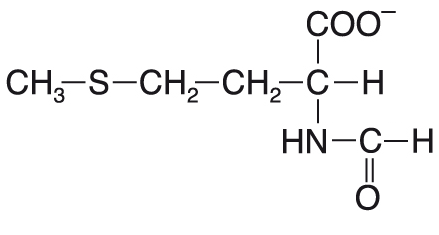 formylací Met-tRNA. formylací Met-tRNA. | |
NGSzkr. sekvenování nové generace, angl. Next-Generation Sequencing, souhrnné označení pro moderní metody sekvenování DNA, zavedené v období od konce 20. století. Tyto nové metody (metody druhé generace) přinášejí oproti klasické Sangerově metodě výhodu v tom, že umožňují rychlejší a levnější sekvenování velkých genomů. DNA je zde rozdělena na krátké úseky, které jsou pak specificky namnoženy a sekvenovány. Při sekvenační reakci katalyzované DNA-polymerasou jsou detekovány jednotlivé komplementární nukleotidy postupně připojované k amplifikovanému vláknu, např. metodami Solid, Solexa nebo Iont Torrent. Tímto způsobem je produkováno velké množství dat (každý fragment je sekvenován několikrát) a získané fragmenty jsou pak skládány pomocí výkonných počítačů do celkové sekvence analyzované DNA. V současnosti jsou k dispozici citlivé metody třetí generace, které umožňují sekvenování jediné molekuly DNA v reálném čase (např. jednomolekulové sekvenování v reálném čase (angl. single-molecule real-time sequencing, SMRT), či nanopórové sekvenování). | |
niacinangl. niacin, viz vitamin B3. | |
ninhydrinangl. ninhydrin, bezbarvá, ve vodě rozpustná sloučenina, velmi často využívaná ke stanovení
aminokyselin. Reakcí s aminoskupinou za zvýšené teploty vzniká fialový produkt. Tímto způsobem reagují všechny volné aminokyseliny; prolin poskytuje žlutý produkt jiné struktury. Ninhydrin reaguje také s volným amoniakem a se všemi primárními aminoskupinami, v bílkovinách a peptidech zejména s N-koncovou aminoskupinou a s ε-aminoskupinou lysinu. 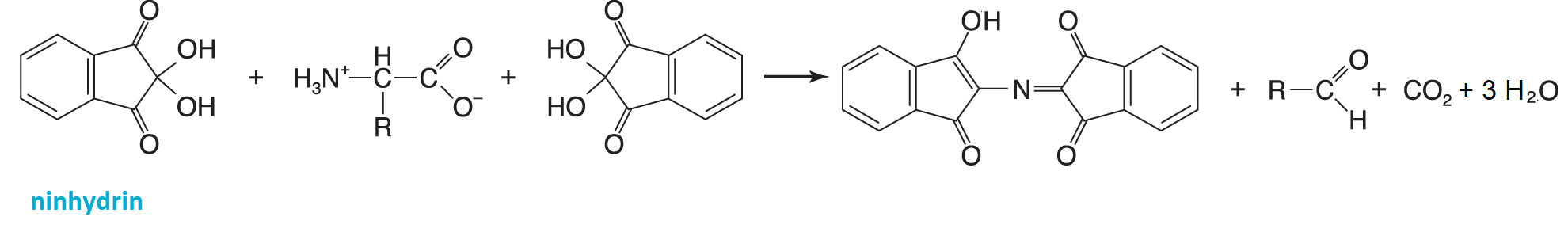 | |
Nirenberg Marshall Warren
| |

nitrátreduktasyangl. nitrate reductases, enzymy schopné redukovat dusičnanový ion na dusitanový (viz koloběh dusíku).
Dissimilační (též respirační) nitrátreduktasa (EC 1.7.5.1, EC 1.9.6.1) je membránový enzym využívající při anaerobní respiraci energii této reakce pro vznik proton-motivní síly. Asimilační nitrátreduktasy (EC 1.7.1.1; EC 1.7.1.2; EC 1.7.1.3, EC 1.7.7.2) se vyskytují u hub a rostlin a umožňují využít nitrátu jako zdroje dusíku pro syntézu aminokyselin. | |
nitrifikaceangl. nitrification, převádění (oxidace) dusíkatých látek na dusičnany (nitráty). Tyto aerobní procesy mají zásadní význam pro
koloběh dusíku v biosféře. Nitrifikační bakterie (viz aerobní chemolithotrofy) mohou pomocí molekulového kyslíku oxidovat amonné ionty (NH4+) na ionty dusitanové (NO2-) nebo dusičnanové (NO3-). | |
nobelisté – přehledNÁSLEDUJE SEZNAM LAUREÁTŮ NOBELOVY CENY, JEJICHŽ OBJEVY ZÁSADNÍM ZPŮSOBEM OVLIVNILY VÝVOJ BIOCHEMIE.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
noradrenalintéž norepinefrin, angl. noradrenaline nebo norepinephrine, patří mezi hormony (působící též jako neurotransmiter) odvozené od tyrosinu (patřící do skupiny katecholaminů, vzorec tamtéž). Vyvolává kontrakci cév (s výjimkou cév srdečních) a zvyšuje tak krevní tlak; uvolňuje naopak hladké svaly, ale stimuluje srdeční sval. | |
Northorp John Howard americký biochemik (1891–1987), Nobelova cena (chemie, 1946) za přípravu enzymů a virových proteinů v čistém stavu (spolu s W. M. Stanleym). Studoval zoologii a chemii na Columbia University. V letech 1917 a 1918 působil jako kapitán chemického vojska. Jeho výzkum byl zaměřen hlavně na sacharidy. Dále se věnoval kinetickým studiím enzymů. V roce 1929 izoloval pepsin v čisté krystalické formě technikami, které později použil se spolupracovníky ke krystalizaci trypsinu, chymotrypsinu, karboxypeptidasy a pepsinogenu. Uspěl při izolaci nukleoproteinu s bakteriofágovou aktivitou. Během II. světové války studoval metody detekce bojových plynů. Ve vědecké práci pak pokračoval studiem virů. americký biochemik (1891–1987), Nobelova cena (chemie, 1946) za přípravu enzymů a virových proteinů v čistém stavu (spolu s W. M. Stanleym). Studoval zoologii a chemii na Columbia University. V letech 1917 a 1918 působil jako kapitán chemického vojska. Jeho výzkum byl zaměřen hlavně na sacharidy. Dále se věnoval kinetickým studiím enzymů. V roce 1929 izoloval pepsin v čisté krystalické formě technikami, které později použil se spolupracovníky ke krystalizaci trypsinu, chymotrypsinu, karboxypeptidasy a pepsinogenu. Uspěl při izolaci nukleoproteinu s bakteriofágovou aktivitou. Během II. světové války studoval metody detekce bojových plynů. Ve vědecké práci pak pokračoval studiem virů. | |
nukleoidtéž bakteriální
chromosom, angl. nucleoid, nukleoproteinový komplex prokaryot tvořený velkou kruhovou molekulou DNA a připojenými bílkovinami. DNA je zde nadšroubovicově svinuta (tvoří superhelixy). Pojem nukleoid se též užívá pro buněčnou oblast, kde je prokaryotní DNA lokalizována. | |
nukleoproteinyangl. nucleoproteins, komplexy
nukleových kyselin s proteiny. Partneři jsou v nich vázáni převážně elektrostatickými interakcemi (nukleové kyseliny jsou záporně nabité, bazické bílkoviny kladně). Typickými představiteli těchto struktur jsou nukleosomy, ribosomy a nukleoidy. | |
nukleosidyangl. nucleosides, glykosidy, jejichž cukernou složku tvoří D-ribosa nebo 2-deoxy-D-ribosa ve furanosové formě, a aglykonem, připojeným 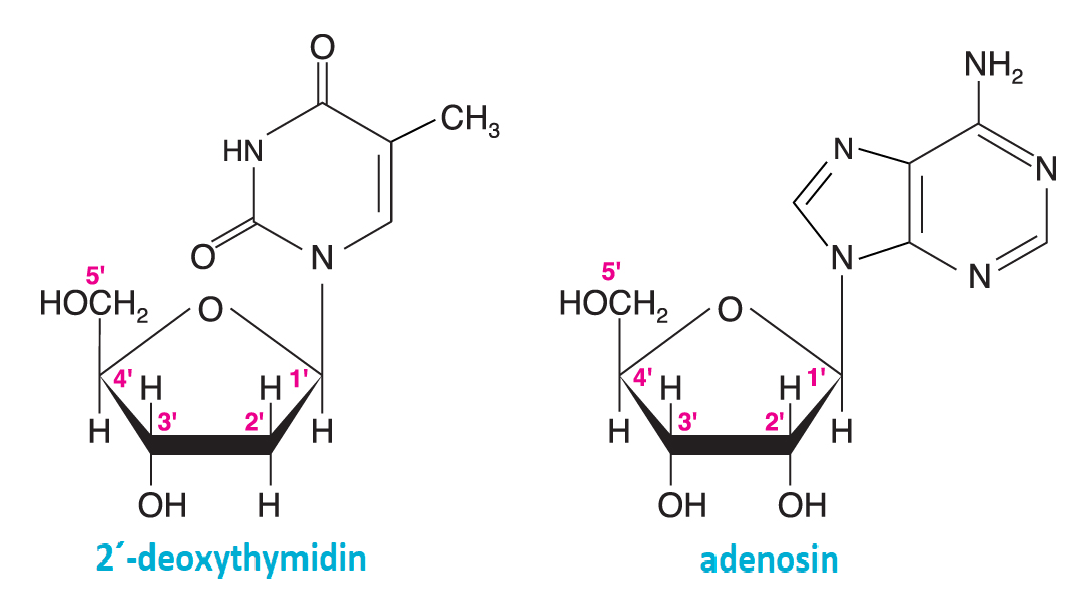 β-N-glykosidovou vazbou, je nukleová báze. Nukleosidy obsahující pyrimidinové báze mají koncovku -idin (cytidin, uridin, deoxythimidin); pokud obsahují purinovou bázi, mají koncovku -osin (adenosin, guanosin). Atomy nukleové báze označujeme běžnými čísly, atomy sacharidu čísly s čárkou; např. správný název cyklický-3′,5′-adenosinmonofosfát (cAMP) ukazuje, že fosfátová skupina je připojena k uhlíku C3 a C5 ribosy. Fosforečné estery nukleosidů nazýváme nukleotidy; nejdůležitější jsou 5′-deriváty; běžné názvosloví pro ně užívá též termíny nukleosid(mono-, di-, tri-)fosfát (např. guanosinmonofosfát, uridindifosfát, cytidintrifosfát, přesněji samozřejmě guanosin-5′-monofosfát atd). β-N-glykosidovou vazbou, je nukleová báze. Nukleosidy obsahující pyrimidinové báze mají koncovku -idin (cytidin, uridin, deoxythimidin); pokud obsahují purinovou bázi, mají koncovku -osin (adenosin, guanosin). Atomy nukleové báze označujeme běžnými čísly, atomy sacharidu čísly s čárkou; např. správný název cyklický-3′,5′-adenosinmonofosfát (cAMP) ukazuje, že fosfátová skupina je připojena k uhlíku C3 a C5 ribosy. Fosforečné estery nukleosidů nazýváme nukleotidy; nejdůležitější jsou 5′-deriváty; běžné názvosloví pro ně užívá též termíny nukleosid(mono-, di-, tri-)fosfát (např. guanosinmonofosfát, uridindifosfát, cytidintrifosfát, přesněji samozřejmě guanosin-5′-monofosfát atd). | |
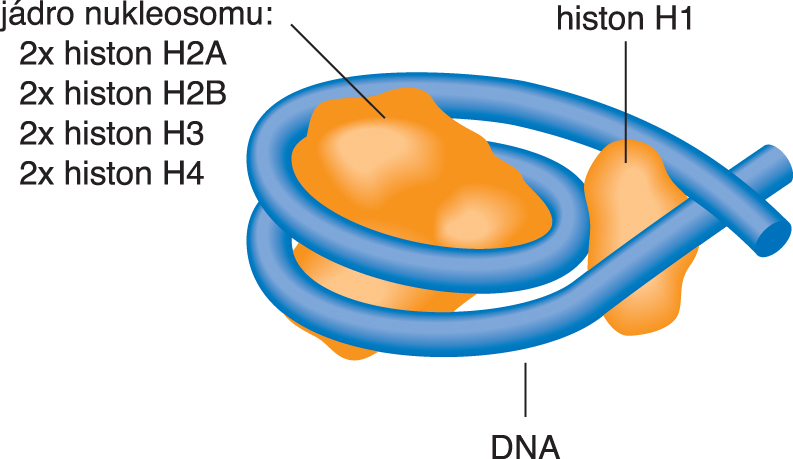
nukleosom146 párů bází (necelé dva závity, jiní autoři udávají 147 párů) navinutých na oktamer histonů. Přibližně 60 následujících párů bází tvoří spojku k dalšímu nukleosomu; někdy se poněkud romanticky hovoří o struktuře eukaryotní DNA jako o "perlovém náhrdelníku". Za rozvoj elektronové mikroskopie krystalů a za určení struktury biologicky významných komplexů nukleových kyselin s bílkovinami (zejména nukleosomů) obdržel roku 1982 A. Klug Nobelovu cenu. | |
nukleotidytéž nukleosidfosfáty, angl. nucleotides, fosforečné estery ribonukleosidů (ribonukleotidy) nebo deoxyribonukleosidů (deoxyribonukleotidy neboli deoxynukleotidy). Fosfátová skupina může být připojena ke kterékoliv volné hydroxylové skupině sacharidu, tedy 2′, 3′ a 5′ u ribonukleotidů a 3′ a 5′ u deoxyribonukleotidů. Mezi nukleotidy řadíme i nukleosidové estery kyseliny difosforečné a trifosforečné (např. ADP, GTP) a také cyklické 3′,5′-fosfodiestery (cAMP, cGMP). Nukleotidy plní v buňkách několik neobyčejně významných funkcí:
| |