Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
B |
|---|
bakterieangl. bacteria, jednobuněčné prokaryotní organismy (viz
organismy – rozdělení podle vnitřní organizace buněk). | |
bakterie denitrifikačníangl. denitrification bacteria, anaerobní chemoorganotrofní bakterie, získávající energii oxidací organických látek. Konečným akceptorem elektronů v dýchacím řetězci je pro ně dusičnanový ion, který se redukuje na dusitan, oxid dusný nebo molekulový dusík (viz anaerobní respirace). Odstraňují nadbytečné dusičnany z biosféry (viz koloběh dusíku). | |
bakterie desulfurikačnítéž desulfurizační, angl. desulphurating nebo desulphurizing microorganisms, anaerobní heterotrofní bakterie, přeměňující při anaerobní respiraci sírany na síru nebo sulfan (SO42- → S → H2S). V hydrogeologických strukturách se uplatňují při redukci síranů ve vodách, hojně se vyskytují v minerálních vodách (např. v Piešťanech). Nacházejí se i ve velkých hloubkách v oceánech. | |
bakterie fototrofnítéž fotosyntetizující, angl. photosynthetic bacteria, zdrojem energie je pro ně světelné záření. Donorem vodíku je u
fotolithotrofních bakterií nejčastěji sulfan (produkují síru), u
fotoorganotrofních bakterií organická látka, např. 2-propanol (produkují
aceton) nebo mastná kyselina (viz anoxygenní fotosyntéza). | |
bakterie halofilníangl. halophilic bacteria nebo haloarcheae, (archea)bakterie, které se dobře rozmnožují při koncentraci chloridu sodného vyšší než 15 %; jsou tedy adaptovány na prostředí s vysokou iontovou silou, a tedy i vysokým osmotickým tlakem. Jsou schopny využívat světlo jako zdroj energie pro vznik proton-motivní síly (bakteriorhodopsin); jde o zvláštní typ fototrofního metabolismu. | |
bakterie hlízkovétéž hlízkovité, angl. nodules-forming bacteria, bakterie rodu Rhizobiaceae, žijící v symbióze na kořenech motýlokvětých rostlin. Mají schopnost redukovat molekulový dusík na amoniak, čímž se podílejí na koloběhu dusíku v přírodě. Rostlině poskytují dusíkaté látky, a naopak od ní přijímají organické látky - produkty fotosyntézy. | |
bakterie methanogenníangl. methanogenous bacteria,
anaerobní chemolithotrofní bakterie (nejčastěji archae). Redukcí oxidu uhličitého
vodíkem (CO2 + 4 H2 → CH4 + 2 H2O) získávají energii, kterou ukládají do ATP
v procesu anaerobní respirace. Žijí ve společenstvích s jinými
typy mikroorganismů, které produkují pro ně potřebný vodík. Využívají se při výrobě
bioplynu. | |
bakterie methanotrofníangl. methanotrophic bacteria, aerobní chemolithotrofní bakterie, jimž jako zdroj energie slouží postupná aerobní oxidace methanu až na oxid uhličitý. Jde o silně exergonický proces; uvažuje se, že by tyto bakterie mohly sloužit jako dodavatelé tepla pro různé technologické procesy. | |
bakterie nitrifikačníangl. nitrifying bacteria, aerobní chemolithotrofy, které prostřednictvím oxidace iontů NH4+ nebo NO2- za vzniku proton-motivní síly získávají ATP. Hrají důležitou roli v koloběhu dusíku v přírodě, protože oxidují amoniak vznikající rozkladem dusíkatých organických sloučenin a produkují dusičnanový ion, který je hlavním zdrojem dusíku pro rostliny. | |
bakterie octovéangl. acetic acid bacteria, bakterie rodu Acetobacter, které oxidují ethanol molekulovým kyslíkem za vzniku octové kyseliny (viz nepravá fermentace). Využívá se jich při výrobě octa, jsou naopak nežádoucí ve vinařství. | |
bakterie sulfurikačnítéž sulfurizační, angl. sulfur-oxidizing bacteria, aerobní chemolithotrofní bakterie, využívající jako zdroj energie oxidaci redukovaných sirných sloučenin (zejména sulfanu) molekulovým kyslíkem. | |
bakterie železitéangl. iron bacteria, aerobní chemolithotrofy, získávající energii oxidací železnatých iontů molekulovým kyslíkem na ionty železité. Často působí problémy tím, že nerozpustnými sloučeninami trojmocného železa ucpávají vodovodní trubky, v nichž proudí voda dostatečně bohatá na železnaté ionty. | |
bakteriofágyangl. bacteriophages, DNA- nebo RNA-viry napadající
bakterie. Mívají velmi vyhraněnou specifitu; napadají jen určité bakteriální druhy. | |
bakteriochlorofylangl. bacteriochlorophyll, pigment fotosyntetizujících bakterií. Ve srovnání s chlorofylem zelených rostlin a řas má absorpční maxima posunuta k delším vlnovým délkám. Proto mohou tyto bakterie žit v oceánech pod vrstvou zeleného planktonu, tvořeného převážně řasami. | |
bakteriorhodopsinangl. bacteriorhodopsin, integrální
membránová bílkovina některých halofilních archeí. Jako prostetickou skupinu obsahuje retinal, který absorbuje viditelné záření; díky této energii je bakteriorhodopsin schopen aktivně transportovat vodíkové nebo sodné ionty, čímž vzniká proton-motivní síla (nebo sodno-motivní síla), kterou ATP-synthasa následně využívá k syntéze ATP. | |
Banting Frederick Grant
| |
 kanadský lékař (1891–1941), Nobelova cena (za fyziologii a lékařství, 1923) za objev
kanadský lékař (1891–1941), Nobelova cena (za fyziologii a lékařství, 1923) za objev barvivaangl. dyes / pigments, v biochemii sekundární metabolity, které absorbují některé složky viditelného světla (a jsou proto barevné). K nejznámějším barvivům rostlin patří flavonoidy, anthokyaniny, karotenoidy a nejdůležitější z nich, chlorofyly. Viz též barviva krevní (hemoglobin) a žlučová. | |
barviva krevníangl. blood pigments, barevné složky krve (červený
hemoglobin, modrý ceruloplasmin, oranžový komplex bilirubinu s albuminem, riboflavin atd.); v užším slova smyslu barviva přenášející kyslík, zejm. hemoglobin. | |
barviva žlučováangl. bile pigments, barevné degradační produkty
porfyrinů, zejména hemu. Ten se postupně přeměňuje v různých buňkách (slezina, játra) na barevné produkty biliverdin, bilirubin a bilirubin-bisglukuronát. Ten je vylučován žlučí do střeva, kde je opět uvolněn bilirubin, který je redukován za přispění střevních bakterií na bezbarvé produkty urobilinogen a sterkobilinogen. Tyto látky jsou kyslíkem oxidovány především na urobilin a sterkobilin, které dodávají stolici charakteristické zbarvení. | |
báze minoritníangl. minor bases, méně často se vyskytující báze nukleových kyselin, modifikace základních bází (methyl-, hydroxy-, thio- nebo hydrogenderiváty). Nacházejí se především v tRNA. Methylace patří k důležitým postsyntetickým modifikacím DNA u prokaryot (viz restrikčně-modifikační systém) i eukaryot (viz epigenetika). Na obr. jsou zachyceny některé často se vyskytující minoritní báze. 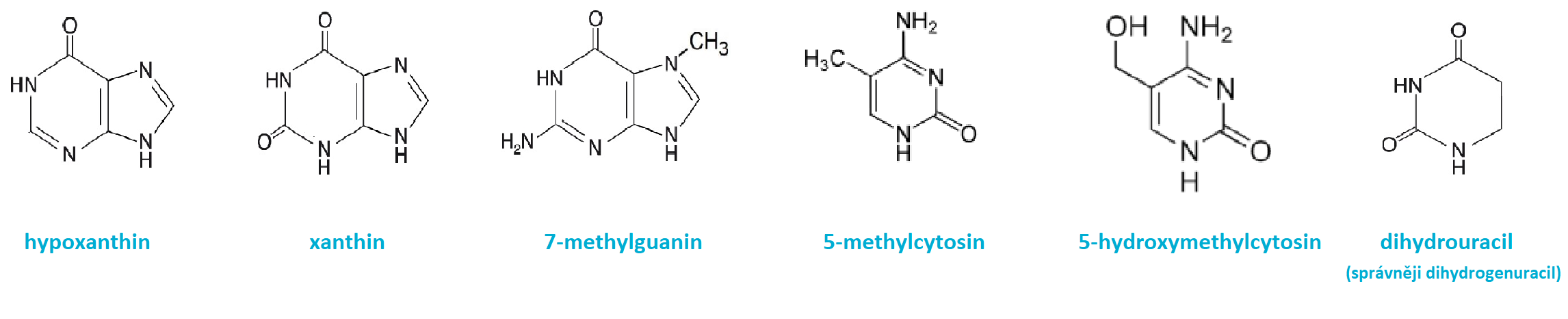 | |
báze nukleových kyselintéž nukleové nebo dusíkaté báze, angl. nucleic acids bases, základní součásti
nukleosidů, nukleotidů i polynukleotidů, tedy nukleových kyselin (báze proto, že mají slabě zásaditý charakter). Dělí se podle heterocyklických sloučenin, které tvoří základ jejich struktury, na báze purinové (adenin, guanin) a pyrimidinové (cytosin, uracil, thymin). Vytvářejí doplňkové dvojice (zvané komplementární páry), v nichž se vždy jedna purinová a jedna pyrimidinová báze vzájemně vážou vodíkovými vazbami . V DNA fungují jako prvky kódu k zápisu genetické informace; jejich komplementarita umožňuje tuto informaci realizovat v procesech replikace, transkripce i translace. | |
Berg Paul
| |

Bergström Sune Karl
| |
 švédský biochemik (1916–2004), Nobelova cena (za fyziologii a lékařství, 1982) za objevy týkající se
švédský biochemik (1916–2004), Nobelova cena (za fyziologii a lékařství, 1982) za objevy týkající se bilance dusíkováangl. nitrogen balance, rozdíl mezi celkovým příjmem dusíku organismem a jeho ztrátou (vylučováním). Mladé rostoucí organismy mají pozitivní dusíkovou bilanci, neboť zadržují dusík, který využívají k výstavbě tkání, zejména ve formě bílkovin. | |
bilirubinangl. bilirubin, žlutooranžová látka, vznikající katabolickým štěpením hemu.  Tvoří se hlavně z hemoglobinu ve slezině, odkud se uvolňuje do krevní plasmy, kde se váže na sérový albumin. Z krve je vychytáván játry a přechází žlučí do střeva, kde je zejména činností bakterií přeměněn na degradační produkty (žlučová barviva). Hromadění bilirubinu v krvi (hyperbilirubinemie) se projevuje žlutým zabarvením kůže, bělimy oka a sliznic (žloutenka neboli ikterus), což je známkou onemocnění jater, ucpání žlučových cest (obstrukční žloutenky) nebo zvýšeným rozpadem červených krvinek (hemolytické anemie). Fyziologicky se žloutenka objevuje u novorozenců, u nichž se po porodu poměrně rychle snižuje počet červených krvinek. V krvi se vyskytuje též ve formě konjugátu s glukuronovou kyselinou (bilirubin-bisglukoronát, nazývaný konjugovaný nebo esterifikovaný bilirubin). Tvoří se hlavně z hemoglobinu ve slezině, odkud se uvolňuje do krevní plasmy, kde se váže na sérový albumin. Z krve je vychytáván játry a přechází žlučí do střeva, kde je zejména činností bakterií přeměněn na degradační produkty (žlučová barviva). Hromadění bilirubinu v krvi (hyperbilirubinemie) se projevuje žlutým zabarvením kůže, bělimy oka a sliznic (žloutenka neboli ikterus), což je známkou onemocnění jater, ucpání žlučových cest (obstrukční žloutenky) nebo zvýšeným rozpadem červených krvinek (hemolytické anemie). Fyziologicky se žloutenka objevuje u novorozenců, u nichž se po porodu poměrně rychle snižuje počet červených krvinek. V krvi se vyskytuje též ve formě konjugátu s glukuronovou kyselinou (bilirubin-bisglukoronát, nazývaný konjugovaný nebo esterifikovaný bilirubin). | |
bílkovinytéž proteiny, angl. proteins, biopolymery, jejichž kostru tvoří polypeptidový řetězec, obsahující obvykle 100 až 2 000 aminokyselinových zbytků. Minimální molární hmotnost bílkovin je 10 000 g/mol; menší oligomery aminokyselin řadíme mezi peptidy. Peptidové řetězce bílkovin jsou syntetizovány na ribosomech procesem zvaným translace; většina z nich podléhá kotranslačním a posttranslačním modifikacím. Pořadí aminokyselin v polypeptidovém řetězci je pro každou bílkovinu jedinečné a geneticky dané. Drobné změny primární struktury (vyvolané mutacemi genu) mohou vést k podstatné změně vlastností dané bílkoviny; této skutečnosti využívá proteinové inženýrství, které se cílenými mutacemi snaží změnit (vylepšit) vlastnosti jednotlivých proteinů. V buňce se vyskytuje několik set až tisíců různých bílkovin, které zajišťují její základní funkce a liší se jak chemickou stavbou (především pořadím aminokyselin v peptidovém řetězci – primární strukturou), tak prostorovým uspořádáním. Studiem závislosti obsahu jednotlivých bílkovin na fyziologickém a patologickém stavu buněk (resp. organismů) se zabývá proteomika. Univerzální systém klasifikace bílkovin neexistuje. Lze je třídit např. na základě rozpustnosti (viz albuminy, globuliny), na základě isoelektrického bodu (kyselé, neutrální, bazické), na základě molekulové hmotnosti (malé do 40 kDa, velké obvykle nad 200 kDa) nebo na základě tvaru molekuly (sferoproteiny, skleroproteiny); zvláštní skupinu tvoří bílkoviny zabudované do biologických membrán (membránové bílkoviny). Podle složení lze bílkoviny dělit na bílkoviny jednoduché a bílkoviny složené; důležitější je však dělení bílkovin podle funkce. Za přípravu enzymů a virových proteinů v čistém stavu získali roku 1946 J. H. Northorp a W. M. Stanley Nobelovu cenu. | |
bílkoviny akutní fázezkratka BAF, angl. acute phase proteins (APP) nebo acute phase reactants (APR), označení proteinů, jejichž koncentrace stoupá o alespoň 25 % během akutní fáze, tedy během zánětu, po vážném zranění nebo během nádorového bujení. Patří k nim zejména C-reaktivní protein (CRP), mannosu-vázající lektin (MBL, mannose-binding lectine), fibrinogen, ceruloplasmin a orosomukoid. Jejich konečným cílem je zvýšit počet bílých krvinek v okolí napadeného místa a bránit tak organismus, dokud nevznikne dostatečné množství protilátek. | |
bílkoviny jednoduchéangl. simple nebo unconjugated proteins, bílkoviny, neobsahující kromě aminokyselinových zbytků, vázaných v polypeptidovém řetězci, žádné další pevně navázané molekuly, skupiny či ionty (srov. bílkoviny složené). | |
bílkoviny membránovéangl. membrane proteins, skupina bílkovin, které 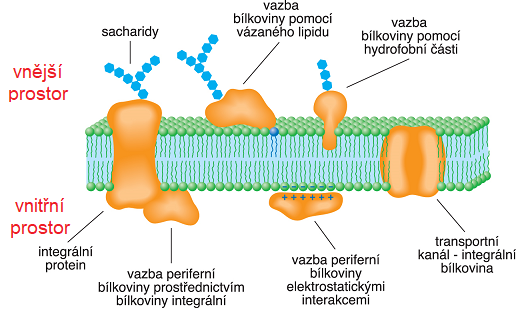 jsou navázány na
biologickou membránu a funkčně s ní souvisejí. Podle síly vazby na dvojvrstvu polárních lipidů je dělíme na periferní, které lze při teplotě 37 °C extrahovat do pufru s nízkou iontovou silou, a integrální, které jsou spojeny s membránou mnohem pevněji. Mezi nejdůležitější transmembránové integrální bílkoviny, jejichž peptidový řetězec prochází (často několikrát) napříč membránou, patří ty, které zajišťují transport částic membránou, dále membránové receptory, odpovídající za interakci buněk s okolím, a membránové enzymy (např. enzymy dýchacího řetězce či světlé fáze fotosyntézy). Mezi periferní bílkoviny patří např. spektrin a aktin, vytvářející v mechanicky silně namáhaných buňkách (nervové buňky, červené krvinky atd.) membránový skelet, který zajišťuje mechanickou odolnost membrány, cytochrom-c, přenášející elektrony v dýchacím řetězci po vnější straně vnitřní mitochondriální membrány, klathrin, hrající významnou roli v endocytose, a G-proteiny, podílející se na přenosu signálu do buňky. jsou navázány na
biologickou membránu a funkčně s ní souvisejí. Podle síly vazby na dvojvrstvu polárních lipidů je dělíme na periferní, které lze při teplotě 37 °C extrahovat do pufru s nízkou iontovou silou, a integrální, které jsou spojeny s membránou mnohem pevněji. Mezi nejdůležitější transmembránové integrální bílkoviny, jejichž peptidový řetězec prochází (často několikrát) napříč membránou, patří ty, které zajišťují transport částic membránou, dále membránové receptory, odpovídající za interakci buněk s okolím, a membránové enzymy (např. enzymy dýchacího řetězce či světlé fáze fotosyntézy). Mezi periferní bílkoviny patří např. spektrin a aktin, vytvářející v mechanicky silně namáhaných buňkách (nervové buňky, červené krvinky atd.) membránový skelet, který zajišťuje mechanickou odolnost membrány, cytochrom-c, přenášející elektrony v dýchacím řetězci po vnější straně vnitřní mitochondriální membrány, klathrin, hrající významnou roli v endocytose, a G-proteiny, podílející se na přenosu signálu do buňky. | |
bílkoviny mléčnéangl. milk proteins, bílkoviny přítomné v mléce. Dělí se na
kaseiny a syrovátkové bílkoviny, z nichž nejdůležitější jsou β-laktoglobulin (tvořící asi 50 % bílkovin syrovátky), α-laktalbumin a laktoferrin. Mléko obsahuje i některé enzymy (např. laktoperoxidasau, EC 1.11.1.7 a xantinoxidasu EC 1.17.3.2). Protilátky z mléka (imunoglobuliny typu IgG, IgA a IgM) jsou přímo (bez štěpení) vstřebávány v tenkém střevu sajícího mláděte a poskytují mu pasivní ochranu proti patogenům, proti nimž je imunní matka. | |
bílkoviny plasmovétéž plasmatické, angl. plasma proteins / plasmatic proteins, skupina bílkovin nacházejících se v krevní plasmě obratlovců (často zúženě chápáno na plasmu člověka). Velká většina z nich jsou složené bílkoviny, především glykoproteiny a lipoproteiny, ale též fosfoproteiny a metaloproteiny. Plasma obsahuje 6 až 8 % proteinů, přičemž zdaleka nejhojnější je sérový albumin (4 až 5 g na 100 ml plasmy). Svými fyzikálně chemickými vlastnostmi plasmové bílkoviny pomáhají regulovat osmotický tlak krve a hodnotu pH. Podílejí se na transportu iontů, hormonů, lipidových látek, vitaminů i některých metabolitů. Jsou odpovědné za koagulaci krve (fibrinogen a koagulační faktory), obranu proti infekcím (imunoglobuliny a systém komplementu) a vykonávají řadu dalších funkcí; některé plní funkci hormonů. Většina je syntetizována v játrech. | |
bílkoviny - podjednotkyangl. protein subunits, jednotlivé nezávisle sbalené polypeptidové řetězce schopné alespoň do jisté míry samostatně vykonávat určité funkce. O podjednotkách hovoříme zejména u bílkovin s kvarterní strukturou. | |
bílkoviny rekombinantníangl. recombinant proteins, proteiny připravené genovými manipulacemi, při nichž byl do genomu producenta uměle začleněn gen kódující požadovanou bílkovinu z jiného organismu (viz transgenní organismy). Tato technologie vyžaduje zvládnutí několika samostatných operací:
| |
bílkoviny – rozdělení podle funkceangl. proteins – functional classification, poněkud násilné, leč velmi užitečné dělení bílkovin; jedna molekula proteinu může totiž současně vykonávat i několik z vyjmenovaných činností. Tímto způsobem obvykle dělíme bílkoviny na:
| |
bílkoviny – rozdělení podle tvaru molekulyangl. proteins – classification according to the molecular shape, klasické dělení proteinů na globulární (též
sferoproteiny) a fibrilární (skleroproteiny). Do tohoto schématu nezapadají membránové bílkoviny. | |
bílkoviny složenétéž konjugované proteiny, angl. conjugated proteins, bílkoviny, obsahující kromě
peptidového řetězce i nepeptidové složky (někdy též méně přesně označované za nebílkovinné). Tak lipoproteiny obsahují lipidy, glykoproteiny obsahují sacharidy, fosfoproteiny zase zbytky kyseliny fosforečné a metaloproteiny obsahují ionty kovů; nukleoproteiny jsou komplexy bílkovin s nukleovými kyselinami. V podstatě terminologicky shodně označujeme složenou bílkovinu jako holoprotein, peptidový řetězec jako apoprotein a nepeptidovou složku jako prostetickou skupinu. | |
bílkoviny strukturníangl. structural proteins, synonymum pro
skleroproteiny. Někdy se tento termín užívá v jiném smyslu pro bílkoviny, které se podílejí na výstavbě struktur určitých orgánů či organel (aktin a myosin ve svalech, některé membránové bílkoviny, bílkoviny spoluvytvářející ribosom apod.). | |
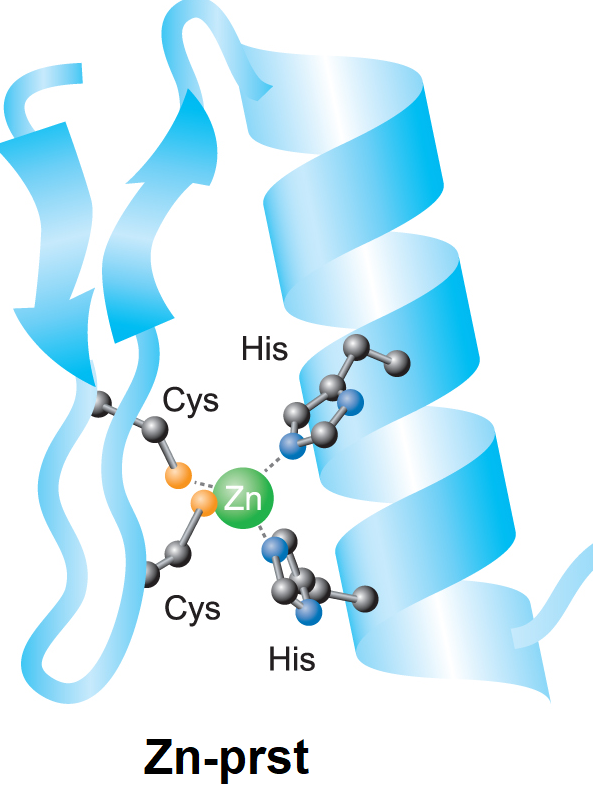
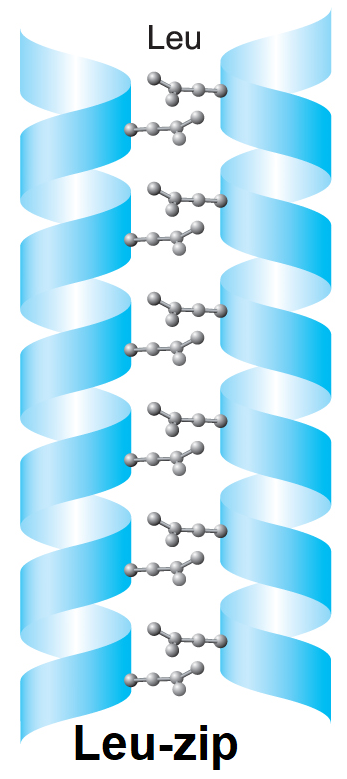
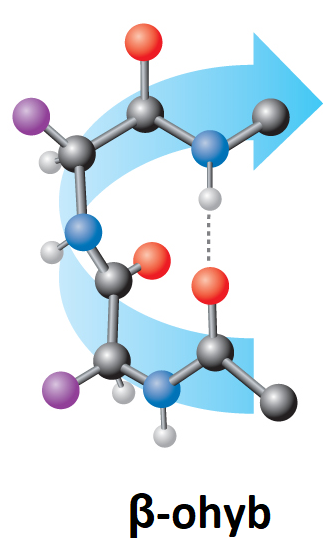
bílkoviny – strukturní motivyangl. proteins – structural motifs, charakteristické prostorové útvary, jež může zaujímat
peptidový řetězec v bílkovinách. Tento modernější pojem postupně vytlačuje starší pojem sekundární struktura. K nejrozšířenějším strukturním motivům patří:
Existenci periodických strukturních motivů bílkovin postuloval L. C. Pauling, který v roce 1954 za výzkum povahy chemické vazby a aplikaci získaných poznatků pro určení struktury komplexních sloučenin získal Nobelovu cenu.
| |
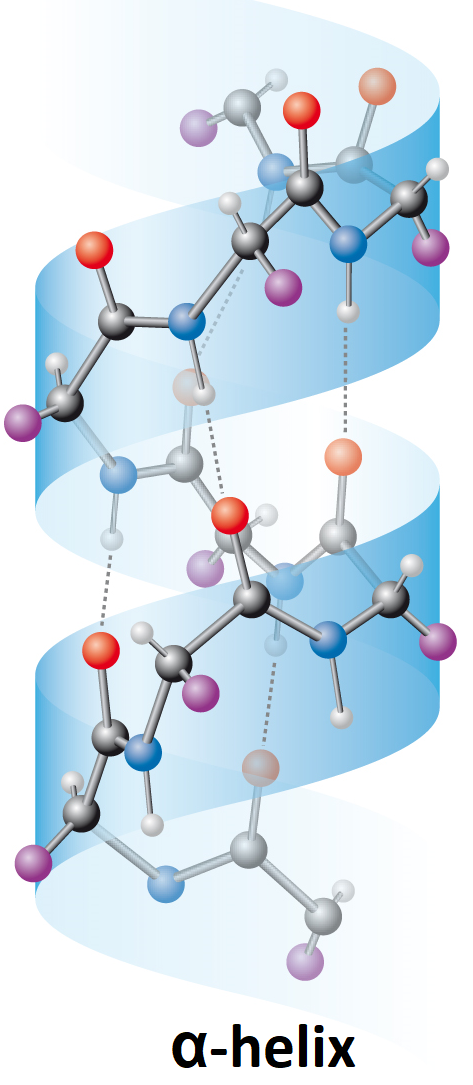
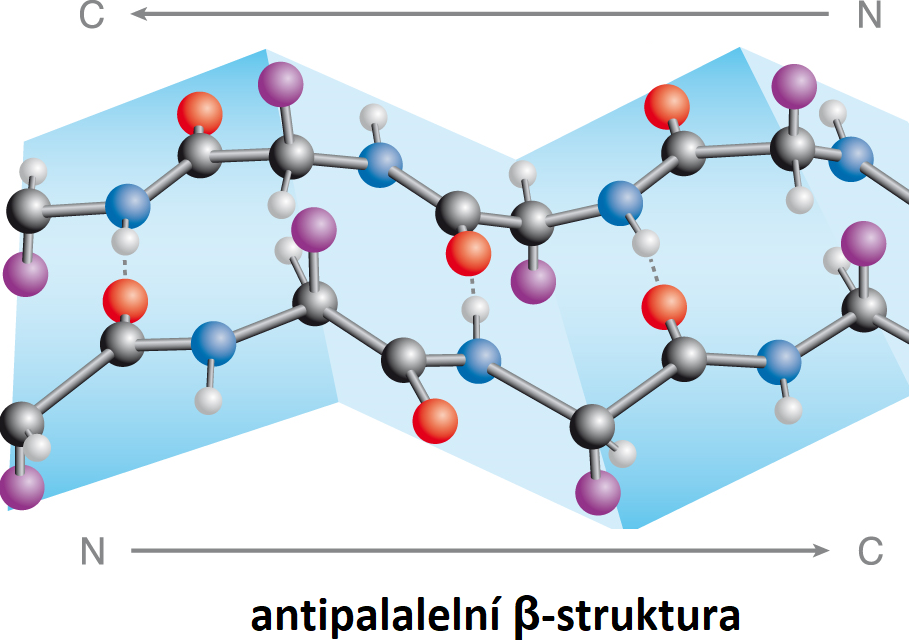
bioenergetikaangl. bioenergetics, interdisciplinární vědní obor na rozhraní mezi
biochemií a fyzikální chemií, zabývající se přeměnami energie v organismech. | |
biochemieangl. biochemistry, vědní obor snažící se vysvětlit biologické děje pomocí chemických prostředků a pojmů. Zabývá se především
| |
biokatalyzátoryangl. biocatalysts, látky, které snižují aktivační energii reakcí a v již nepatrných množstvích ovlivňují různé životní procesy. Hlavní skupinu biokatalyzátorů tvoří:
| |
bioluminiscenceangl. bioluminiscence, emise světla některými organismy (světlušky, medúzy, mnohé hlubokomořské ryby). Světlo je emitováno jako výsledek oxidace druhově specifických látek (alkoholů, aldehydů apod.) molekulovým kyslíkem; tyto látky se nazývají
luciferiny a enzymy, které jejich oxidaci katalyzují, jsou luciferasy. Produkt reakce je v elektronově excitovaném stavu a samovolně přechází do základního stavu zářivým přechodem (emise záření). Během tohoto procesu se velká část reakční energie (až 95 %) uvolní jako viditelné záření (světlo). Pro svou citlivost se bioluminiscence využívá k laboratornímu stanovením některých substrátů, velmi často ATP. | |
biomakromolekulytéž biopolymery, angl. biomacromolecules nebo biopolymers, makromolekuly, vyskytující se v organismech. Podle tradiční definice mezi ně patří látky, jejichž molekulová hmotnost přesahuje 10 kDa, tedy zejména bílkoviny, nukleové kyseliny a polysacharidy. | |
biopolymeryangl. biopolymers, souhrnný název pro
bílkoviny, nukleové kyseliny a polysacharidy (viz struktura biopolymeru kovalentní, nativní, kooperativní, neuspořádaná, struktura biopolymeru primární, prostorová, struktura biopolymeru sekundární, neperiodická, supersekundární, terciární, kvarterní). | |
bioreaktoryangl. bioreactors, obecné označení pro technologická zařízení, ve kterých probíhá chemická reakce v důsledku působení
enzymů nebo častěji živých buněk (mikroorganismú, rostlinných buněk atd., zvaná fermentory). Používají se např. při fermentačních výrobách a při čištění odpadních vod biologickou cestou. Dělí se na aerobní (obvykle probublávané vzduchem) a anaerobní, v nichž probíhají děje bez přístupu kyslíku, dále pak podle technického provedení na míchané a nemíchané, otevřené a uzavřené atd. | |
biosenzoryangl. biosensors, analytická zařízení obsahující biologickou součást (např.
enzym, protilátku nebo i imobilizované buňky). Interakcí analytu s biologickou komponentou je generován elektrický nebo optický signál, který je následně zesílen a dále zpracován (viz elektrody enzymové). | |
biosyntézaangl. biosynthesis, syntetický proces probíhající v organismu. Soubor těchto procesů (též drah) nazýváme
anabolismus. | |
biosyntéza isoprenoidůangl. isoprenoid biosynthesis, proces vycházející z acetyl-CoA, při němž vzniká základní pětiuhlíkatý aktivovaný prekursor biosyntézy všech isoprenoidů, isopentenyldifosfát:
3 CH3–CO–SCoA + 2 (NADPH + H+) + 3 ATP + 2 H2O → CH2=C(CH3)–CH2–CH2–O–PP + CO2 + 3 ADP + Pi + 2 NADP+ + 3 HSCoA Opakovanou kondenzací isopentenyldifosfátu a jeho isomeru, dimethylallyldifosfátu, provázenou odštěpením PPi, vznikají kostry všech isoprenoidních struktur, které pak mohou procházet dalšími úpravami (cyklizacemi, hydroxylacemi, hydrogenacemi apod.). | |
biosyntéza mastných kyselinangl. fatty acids biosynthesis, děj probíhající v mnoha tkáních (např. v játrech, ledvinách, mozku, plících, v tukových tkáních, mléčné žláze). Primárním
prekursorem syntézy je acetyl-CoA, který je nejdříve aktivován karboxylací za vzniku malonyl-CoA. Ten v následujícím kroku kondenzuje za odštěpení oxidu uhličitého s již existujícím acylovým zbytkem, vzniká 3-oxoacylový zbytek, který je v několika krocích redukován za vzniku nasyceného řetězce. Tento v podstatě cyklický děj se několikrát opakuje; vznikají tak mastné kyseliny se sudým počtem uhlíků:
Postupně se prodlužující acylový zbytek je vázán na zvláštní protein (ACP), který je u živočichů součástí multifunkčního enzymu synthasa mastných kyselin (EC 2.3.1.85), katalyzujícího celý popisovaný sled reakcí. Biosyntéza mastných kyselin je regulována v prvním kroku, tedy karboxylaci acetyl-CoA, hormonálně prostřednictvím allosterických efektorů a kovalentních modifikací enzymu. | |
biosyntéza purinových nukleotidů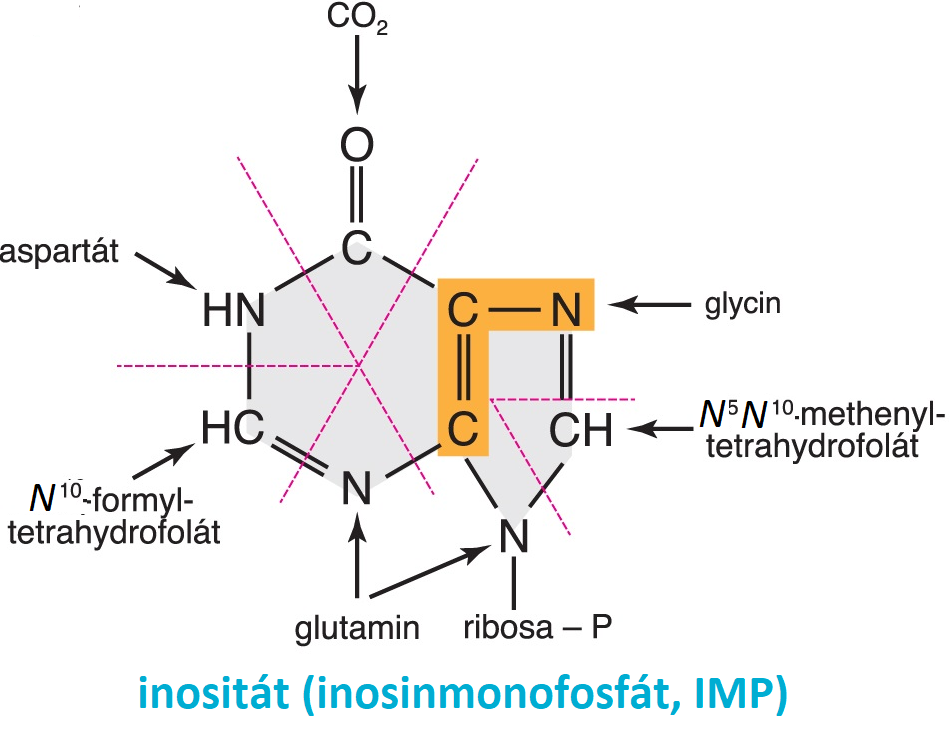 angl. purine nucleotides biosyntesis, proces zahájený vznikem 5-fosforibosyl-1-difosfátu, k němuž se postupně připojují malé části purinového skeletu: dva dusíky z glutaminu, molekula glycinu, pak methyl z tetrahydrofolátového koenzymu, uhlík z oxidu uhličitého, dusík z asparagové kyseliny a uhlík z tetrahydrofolátového koenzymu; purinové jádro tak vzniká jako mozaika atomů pocházejících z různých molekul. angl. purine nucleotides biosyntesis, proces zahájený vznikem 5-fosforibosyl-1-difosfátu, k němuž se postupně připojují malé části purinového skeletu: dva dusíky z glutaminu, molekula glycinu, pak methyl z tetrahydrofolátového koenzymu, uhlík z oxidu uhličitého, dusík z asparagové kyseliny a uhlík z tetrahydrofolátového koenzymu; purinové jádro tak vzniká jako mozaika atomů pocházejících z různých molekul. | |
biosyntéza pyrimidinových nukleotidůangl. pyrimidine nucleotides biosynthesis, proces, který vychází z karbamoylfosfátu (oranžová), 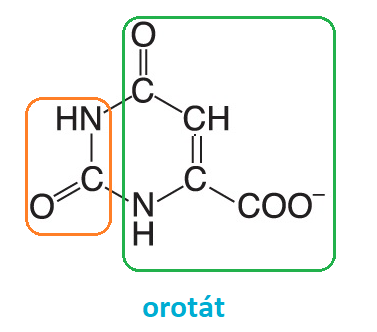 který kondenzuje s asparagovou kyselinou (zelená) za vzniku látek, které již mají základní strukturu pyrimidinového heterocyklu (srov. postupnou výstavbu purinového jádra). Nejdůležitější z těchto derivátů je orotát, který reaguje s 5-fosforibosyl-1-difosfátem za vzniku nukleotidu, jenž je dále upravován na nukleotidy základních bází. který kondenzuje s asparagovou kyselinou (zelená) za vzniku látek, které již mají základní strukturu pyrimidinového heterocyklu (srov. postupnou výstavbu purinového jádra). Nejdůležitější z těchto derivátů je orotát, který reaguje s 5-fosforibosyl-1-difosfátem za vzniku nukleotidu, jenž je dále upravován na nukleotidy základních bází. | |
biosyntéza triacylglycerolů a fosfatidůtéž lipogenese, angl. triacylglycerol and phosphatide biosynthesis, probíhá v cytosolu, 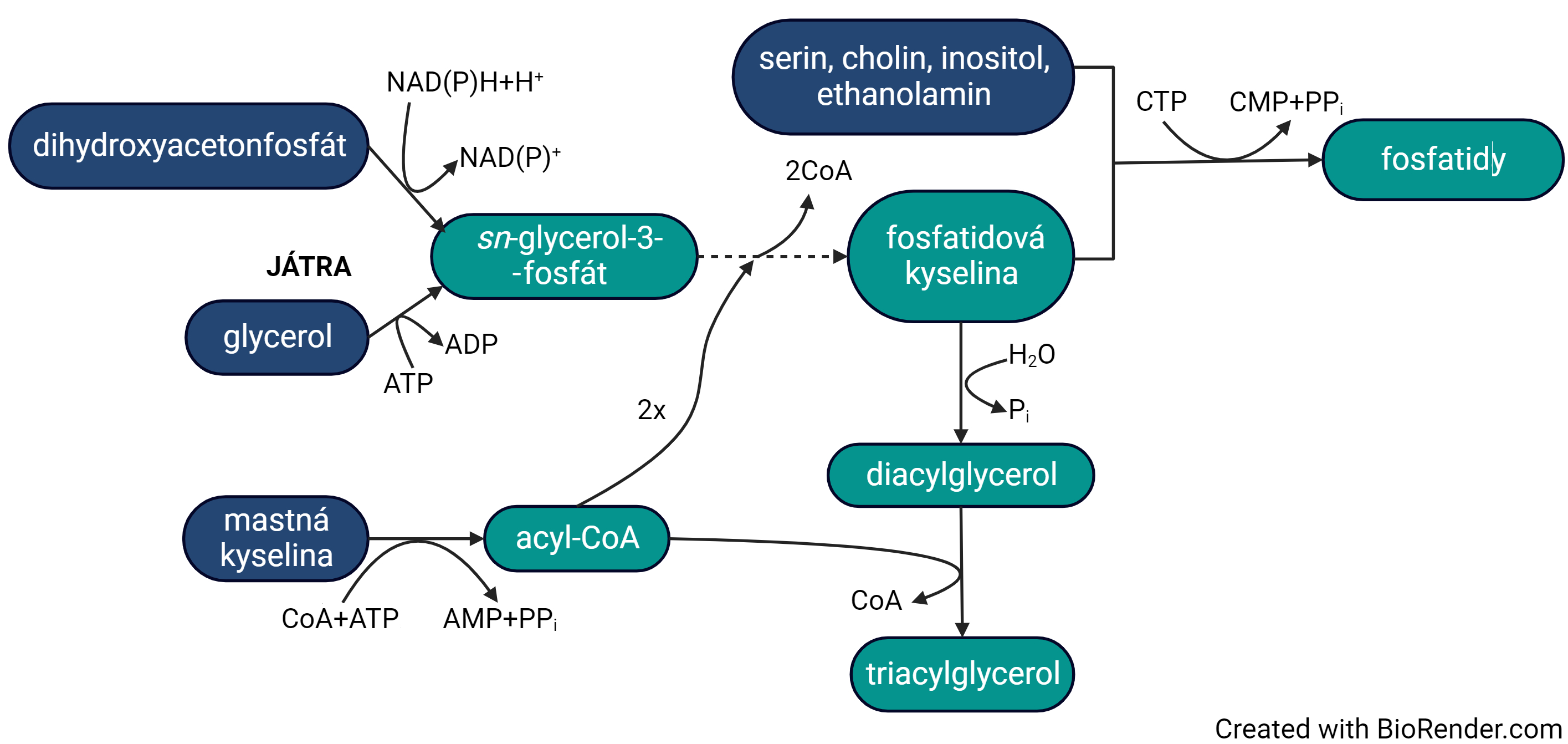 u obratlovců především v jaterních a v tukových buňkách. Vychází z aktivovaných mastných kyselin v podobě acyl-CoA, které jsou transferasami přenášeny na sn-glycerol-3-fosfát (aktivovaný glycerol vznikající buď redukcí dihydroxyacetonfosfátu nebo fosforylací glycerolu); tímto způsobem vzniká fosfatidová kyselina. Z ní mohou být za současného štěpení CTP syntetizovány fosfatidy, nebo po hydrolytickém odštěpení fosfátu může být další transferasovou reakcí připojen třetí acylový zbytek a vznikne triacylglycerol. u obratlovců především v jaterních a v tukových buňkách. Vychází z aktivovaných mastných kyselin v podobě acyl-CoA, které jsou transferasami přenášeny na sn-glycerol-3-fosfát (aktivovaný glycerol vznikající buď redukcí dihydroxyacetonfosfátu nebo fosforylací glycerolu); tímto způsobem vzniká fosfatidová kyselina. Z ní mohou být za současného štěpení CTP syntetizovány fosfatidy, nebo po hydrolytickém odštěpení fosfátu může být další transferasovou reakcí připojen třetí acylový zbytek a vznikne triacylglycerol. | |
biotechnologieangl. biotechnology, obor, využívající organismů, buněk, jejich součástí a molekulárních analogů k výrobě produktů a ke službám. Nejstaršími biotechnologiemi, známými již ve starověku, jsou kvasné procesy (viz fermentace). V moderním pojetí jde hlavně o postupy založené na znalostech molekulové genetiky s využitím genového inženýrství. Patří sem např. produkce antibiotik fermentačními postupy, výroba rekombinantních bílkovin, koncentrování minerálů pomocí mikroorganismů (loužení rud), příprava biosenzorů pro analytickou chemii, výroba průmyslových enzymů (pro prací prášky apod.), degradace odpadů mikroorganismy (např. čištění půdy zamořené ropnými produkty, bioplynové stanice), biotransformace některých látek a produkce monoklonálních protilátek. | |
biotinangl. biotin, viz vitamin B7. | |
biotransformace cizorodých látekangl. xenobiotic transformation, součást procesu vylučování cizorodých látek (xenobiotik), zejména farmak a toxických látek, z organismu. Některá xenobiotika se sice mohou z organismu vyloučit beze změny, většina však musí být transformována: v první fázi se obvykle hydroxylují a ve druhé se navážou na polární metabolity (glukuronová kyselina, glutathion, některé aminokyseliny). Tím se zpravidla zvýší jejich rozpustnost ve vodě, a tak se usnadní jejich vyloučení. Jinou častou biotransformací je methylace. Studiem biotransformací se zabývá xenobiochemie. | |
biotransformace průmyslovátéž biokonverze, angl. industrial biotransformation, obvykle jednostupňová chemická přeměna určité látky realizovaná pomocí organismů (většinou mikroorganismů) nebo jejich součástí (enzymů, viz biotechnologie). Biotransformacemi se běžně vyrábějí např. octová kyselina (z ethanolu), sorbosa (pro produkci askorbové kyseliny), některé steroidy, salicylová kyselina a 6-aminopenicilinová kyselina. | |
bisfosfoglycerátangl. bisphosphoglycerate, pod tento termín se zahrnuje:
| |
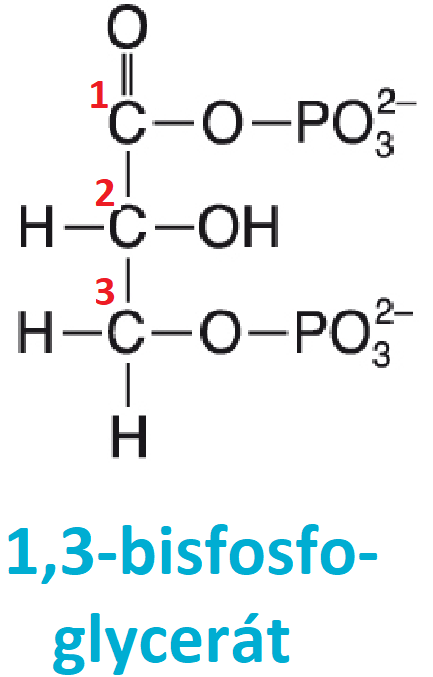 V
V Blackburn Elizabeth Helen
| |
 molekulární bioložka (nar. 1948), Nobelova cena (za fyziologii a lékařství, 2009) za příspěvek k pochopení funkce
molekulární bioložka (nar. 1948), Nobelova cena (za fyziologii a lékařství, 2009) za příspěvek k pochopení funkce Blobel Günter
| |

Bloch Konrad Emil
| |

bod iniciačnítéž bod replikační, angl. iniciation point nebo replication origin, počáteční bod
replikace DNA. Úsek, replikovaný z jednoho iniciačního bodu, se nazývá replikon. Chromosomy prokaryotních buněk, plasmidy a virová DNA se replikují z jediného iniciačního bodu (tvoří jediný replikon); chromosomy eukaryotní buňky obsahují asi 1000 replikonů, z nichž každý má délku přes 10 μm. | |
bod isoelektrickýangl. isoelectric point, zkr. pI, hodnota pH roztoku, při které (pH = pI) se
amfion nepohybuje v elektrickém poli; to znamená, že jeho volný náboj je zde nulový. Isoelektrický bod lze určit pro každý amfion, tedy zejména pro aminokyseliny, peptidy a bílkoviny. Jeho hodnota (zejména u bílkovin) výrazně závisí na složení pufru, v němž se provádí elektroforéza (srov. isoionický bod); proto je nutné toto složení pufru vždy uvádět spolu s experimentálně stanovenou hodnotou pI. | |
bod isoionickýangl. isoionic point, hodnota pH, při němž má
amfion, rozpuštěný v destilované vodě, nulový náboj. Hodnota isoionického bodu poskytuje lepší informaci o iontových vlastnostech amfiontu než hodnota bodu isoelektrického, je však obtížnější ho experimentálně stanovit. | |
Boyer Paul Delos
| |

Brown Michael Stuart
| |

bublina replikačnítéž replikační očko, angl. replication bubble, úsek 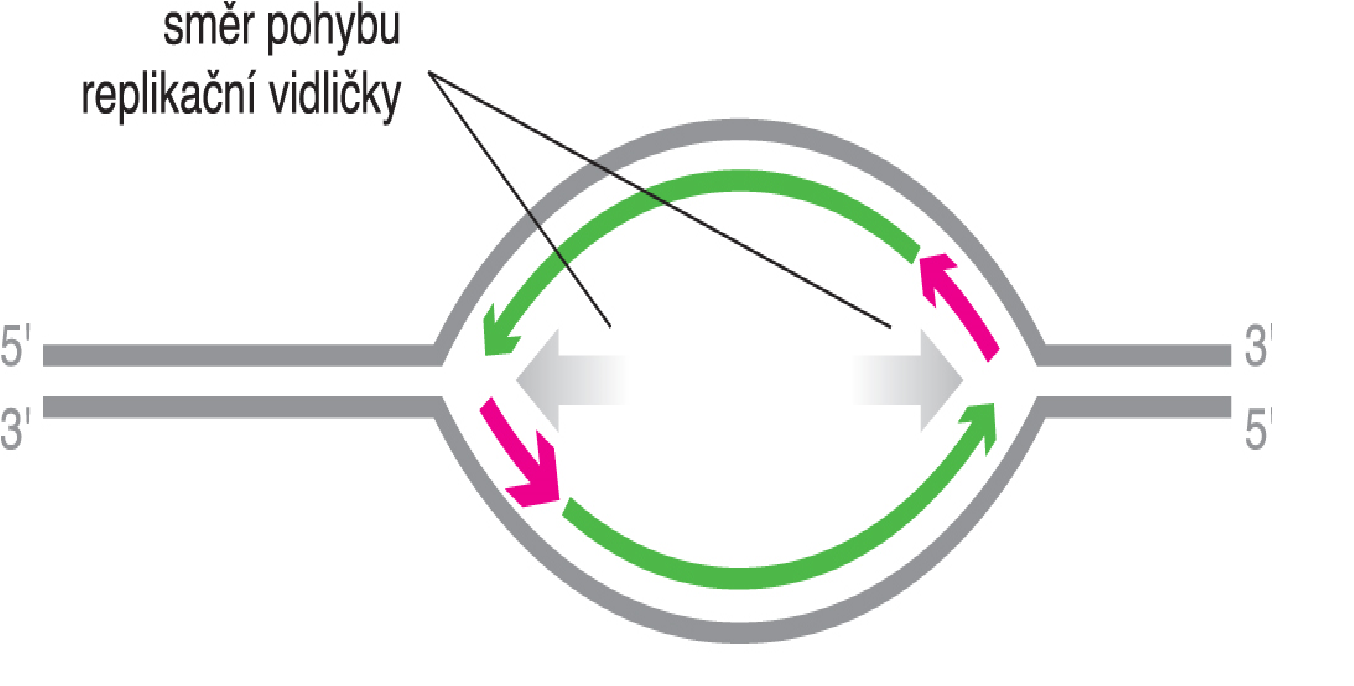 dvouřetězcové
DNA, v němž byly přerušeny vodíkové můstky a řetězce se oddálily tak, aby na nich mohla začít replikace. dvouřetězcové
DNA, v němž byly přerušeny vodíkové můstky a řetězce se oddálily tak, aby na nich mohla začít replikace. | |
Buchner Eduard
| |
 německý chemik a enzymolog (1860–1917), Nobelova cena (chemie, 1907) za objev bezbuněčného
německý chemik a enzymolog (1860–1917), Nobelova cena (chemie, 1907) za objev bezbuněčného bujení rakovinnétéž maligní růst nebo nádorové bujení, angl. cancer proliferation, nekontrolovaný růst a dělení buněk způsobené ztrátou kontroly organismu nad
rozhodujícími molekulárně genetickými procesy. Některé rakovinné buňky mají schopnost migrovat a vytvářet sekundární nádory (metastázy). | |
buňkaangl. cell, základní organizační jednotka živé hmoty. Hlavní myšlenky současné buněčné teorie života lze shrnout do následujících bodů:
1. Buňky tvoří veškerou živou hmotu. 2. Veškeré buňky pocházejí z buněk (dříve existujících, mateřských). 3. Genetické informace se předávají z jedné generace buněk na další. 4. V buňkách látky podléhají chemickým přeměnám. 5. Buňky reagují na vnější podněty. Strukturně se buňky navzájem velmi liší, jsou však všechny ohraničeny buněčnou membránou, základ jejich obsahu tvoří vysoce koncentrovaný roztok bílkovin (cytosol) a obsahují genetickou informaci uloženou v molekule DNA. Podle vnitřní organizace buněk dělíme organismy na prokaryota a eukaryota. | |
buňky diploidníangl. diploid cells, buňky obsahující ve své genetické výbavě každý
chromosom ve dvou homologních kopiích (vyjma chromosomů pohlavních, které mohou být přítomny pouze v jediné kopii, srov. buňky haploidní). | |
buňky haploidníangl. haploid cells, buňky, které ve své genetické výbavě obsahují každý
chromosom pouze v jediné kopii (pohlavní buňky), srov. buňky diploidní. | |
buňky kmenovéangl. stem cells, málo diferencované nebo vůbec nediferencované buňky, z nichž se mohou diferenciací vyvinout linie funkčních diferencovaných buněk. Kmenovými buňkami podle této definice jsou např. buňky zárodku (zygoty a blastuly) nebo pluripotentní kmenové buňky kostní dřeně, z nichž se vyvíjejí všechny druhy krevních buněk. V současné době je snaha použít kmenové buňky jako náhradu tělních buněk, které jsou nějakým způsobem poškozené, nebo z nich vypěstovat
náhradníorgány. Snahy používat kmenové buňky pro účely reprodukční medicíny se považují z etických důvodů za nepřípustné. | |
buňky mnohojadernéangl. polynucleate cells, buňky, které vznikly splynutím (viz buněčná fúze) několika buněk stejného typu, přičemž jejich jádra zůstala zachována (např. buňky příčně pruhovaného svalu). | |
buňky pohlavnítéž gamety, angl. sex cells, buňky s polovičním počtem chromosomů (haploidní), vzniklé
meiózou. Dělíme je na samičí (vajíčka u živočichů, oosféra u rostlin) a samčí (spermie u živočichů, spermatické buňky v pylu vyšších rostlin). | |
buňky polyploidníangl. polyploid cells, buňky s větším počtem sad chromosomů než dvě (srov. diploidní buňky), u živočichů obvykle s menší vitalitou. U rostlin se využívají k šlechtění. Některé odrůdy jsou např. tetraploidní nebo hexaploidní, liché stupně ploidie (tri-, penta-...) tvoří obvykle sterilní organismy (viz též buňky mnohojaderné). | |
Butenandt Adolf Friedrich Johan
| |