Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
R |
|---|
racemasyangl. racemases, enzymy ze třídy
isomeras (EC 5.1.-.-), které katalyzují změnu konfigurace chirálních látek s jedním chirálním centrem (srov. epimery). V metabolismu mikroorganismů se uplatňuje řada racemas aminokyselin. | |
racemizaceangl. racemisation, proces, při němž se samovolně nebo katalyticky (viz
racemasy) mění konfigurace stereogenního centra. Z původní „opticky čisté“ sloučeniny (např. L-Ala) získáme racemickou směs, tvořenou oběma enantiomery (D-Ala i L-Ala); optická rotace tak postupně klesá k nule. | |
radiace fotosynteticky aktivníangl. photosynthetically active radiation, PAR, oblast záření aktivního při
fotosyntéze určitého organismu. Pro rostlinné buňky je to oblast viditelného záření (400 až 500 nm a 600 až 700 nm), fototrofní bakterie využívají záření až do vlnových délek 1 000 nm. Absorpce záření určité vlnové délky fototrofním organismem je závislá na struktuře a obsahu fotosyntetických pigmentů a na anatomické struktuře fotosyntetického orgánu (nejčastěji listu). | |
radikál hydroxylovýOH•, angl. hydroxyl radical, jedna z reaktivních kyslíkových částic, způsobujících oxidační stres. Vzniká především jednoelektronovou redukcí peroxidu vodíku: H2O2 + e− + H+→ OH• + H2O. | |
radikál superoxidovýO2−•, angl. superoxide radical, jedna z reaktivních kyslíkových částic způsobujících oxidační stres. Vzniká jednoelektronovou redukcí molekulového kyslíku. Je odstraňován enzymem superoxiddismutasou. | |
radioimunoanalýzaangl. radioimmunoassay, RIA, imunochemická technika, při níž jsou
protilátka nebo antigen značeny radionuklidem. Užívá se zejména pro stanovení některých hormonů. Do vzorku (obvykle krevního séra) se přidá jisté množství radioaktivně označeného hormonu. Směs se nechá reagovat s protilátkou proti stanovovanému hormonu; protilátky musí být v tomto uspořádání (nazývá se kompetitivní) stechiometricky méně než celkového množství hormonu. Komplex protilátky s navázaným hormonem se separuje (obvykle je protilátka adsorbována na stěny reakční nádobky); čím vyšší je jeho radioaktivita, tím méně bylo ve vzorku daného hormonu. V současné době popularita radioimunoanalýzy klesá, neboť laboratoře se snaží vyhnout práci s radioaktivními izotopy. Za vývoj metod radioimunoanalýzy pro stanovení peptidových hormonů obdržela roku 1977 R. S. Yalow Nobelovu cenu. | |
rafty membránovétéž membránové mikrodomény, angl. membrane rafts (raft = vor), uspořádané oblasti v biologické membráně eukaryotních buněk. Obsahují cholesterol, sfingolipidy a příslušné bílkoviny a zajišťují některé specifické funkce. Lipidy v těchto oblastech tedy mají omezenou laterální pohyblivost, membrány jsou zde silnější a tužší. | |
Ramakrishnan Venkatraman
| |

rámec čtecí | |
rámec čtecí otevřenýangl. open reading frame, ORF, deoxyribonukleotidová sekvence ohraničená z jedné strany kodonem pro methionin (nebo jiným iniciačním kodonem) a z druhé strany STOP-kodonem; pojem užívaný při in silico analýze genomové DNA u prokaryot nebo cDNA u eukaryot. Každou nukleotidovou sekvenci lze přeložit do šesti různých aminokyselinových sekvencí (tři v jednom směru vždy posun začátku čtení tripletů o jednu bázi a tři analogicky v druhém směru) a v každé z nich můžeme ORF nalézt. Porovnáním získaných variant ORF s databázemi můžeme identifikovat, zda daný ORF představuje gen kódující protein. Na obrázku je příklad překladu ve všech šesti čtecích rámcích s vyznačením sekvencí odpovídajících otevřeným čtecím rámcům (červeně), přičemž za sekvenci kódující protein lze (díky její délce) považovat pouze červený úsek v horní části obrázku (5′-3′-Frame 1).  | |
reakce endergonickétéž procesy endergonické, angl. endergonic reactions nebo endergonic processes, děje, které jsou (samy o sobě) za fyziologických podmínek energeticky nevýhodné (změna Gibbsovy energie je pozitivní,
ΔG > 0) a mohou tedy probíhat pouze v případě, že jsou bezprostředně spřaženy s reakcí (dějem), který jim energii dodává. Typickým příkladem jsou reakce katalyzované ligasami, kdy syntetická reakce by nemohla probíhat, kdyby nebyla spřažena se štěpením makroergické sloučeniny (obvykle ATP). Endergonickými ději jsou též např. aktivní transport, na GTP závislá vazba aminoacyl-tRNA na ribosom (viz translace) nebo membránová fosforylace, kde endergonická syntéza ATP je umožněna energií poskytnutou pasivním transportem H+ (srov. reakce exergonické). | |
reakce exergonickétéž procesy exergonické, angl. exergonic reactions nebo exergonic processes, děje, které jsou za fyziologických podmínek doprovázeny poklesem Gibbsovy energie (
ΔG < 0) a mohou tedy z termodynamického hlediska probíhat samovolně (srov. endergonické reakce). | |
reakce polymerasová řetězováangl. polymerase chain reaction, PCR, technika umožňující získat velké množství kopií určité sekvence
DNA. Cyklicky se opakuje teplotní denaturace (t ≈ 95 °C), při níž se rozpadá dvouřetězcová DNA na samostatné řetězce, a replikace, při níž vzniklé jednotlivé řetězce DNA slouží jako templáty pro DNA-polymerasu (t ≈ 70 °C). Pro iniciaci replikace musí být nejdříve připojeny primery (t ≈ 50 °C); jejich struktura rozhoduje o tom, kde replikace začne a který úsek DNA tedy bude namnožen. Počet kopií tohoto úseku roste exponenciálně s počtem cyklů (proto "řetězová rekce"). Za objev polymerasové řetězové reakce získal K. B. Mullis roku 1993 Nobelovu cenu. 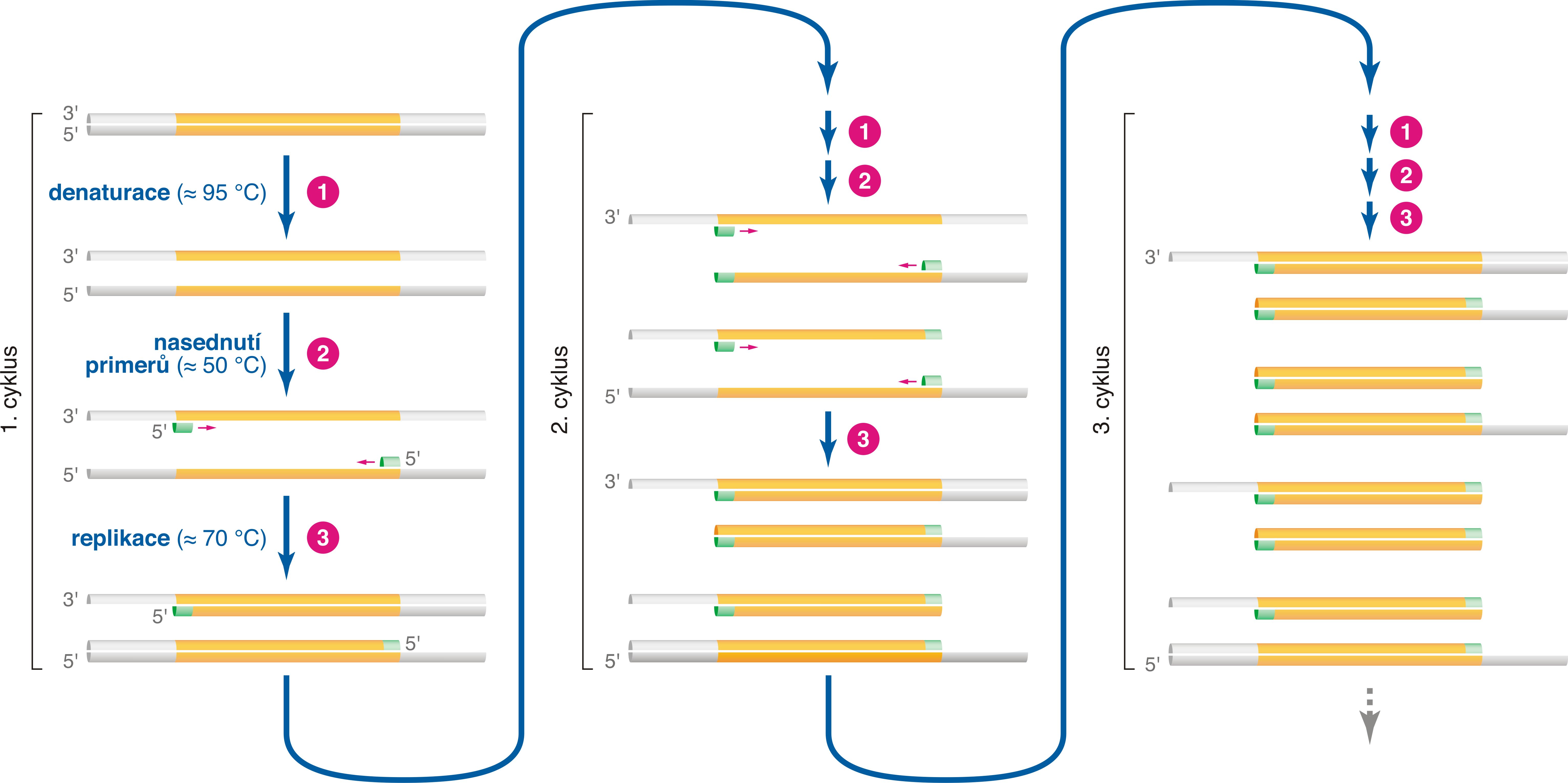 | |
receptor tyrosinkinasovýangl. receptor tyrosin kinases (RTKs), skupina receptorů s vlastní tyrosinkinasovou aktivitou. Nacházejí se v buněčné membráně. Po vazbě ligandu (například insulinu) na tento receptor dojde k aktivaci tyrosinkinasové domény receptoru uvnitř buňky a fosforylaci tyrosinových zbytků v této části molekuly. Takto aktivovaný receptor pak může fosforylovat další intracelulární enzymy, které se tak aktivují a spouštějí kaskádu reakcí v odpovědi na vazbu ligandu na receptor (viz transdukce signálu). 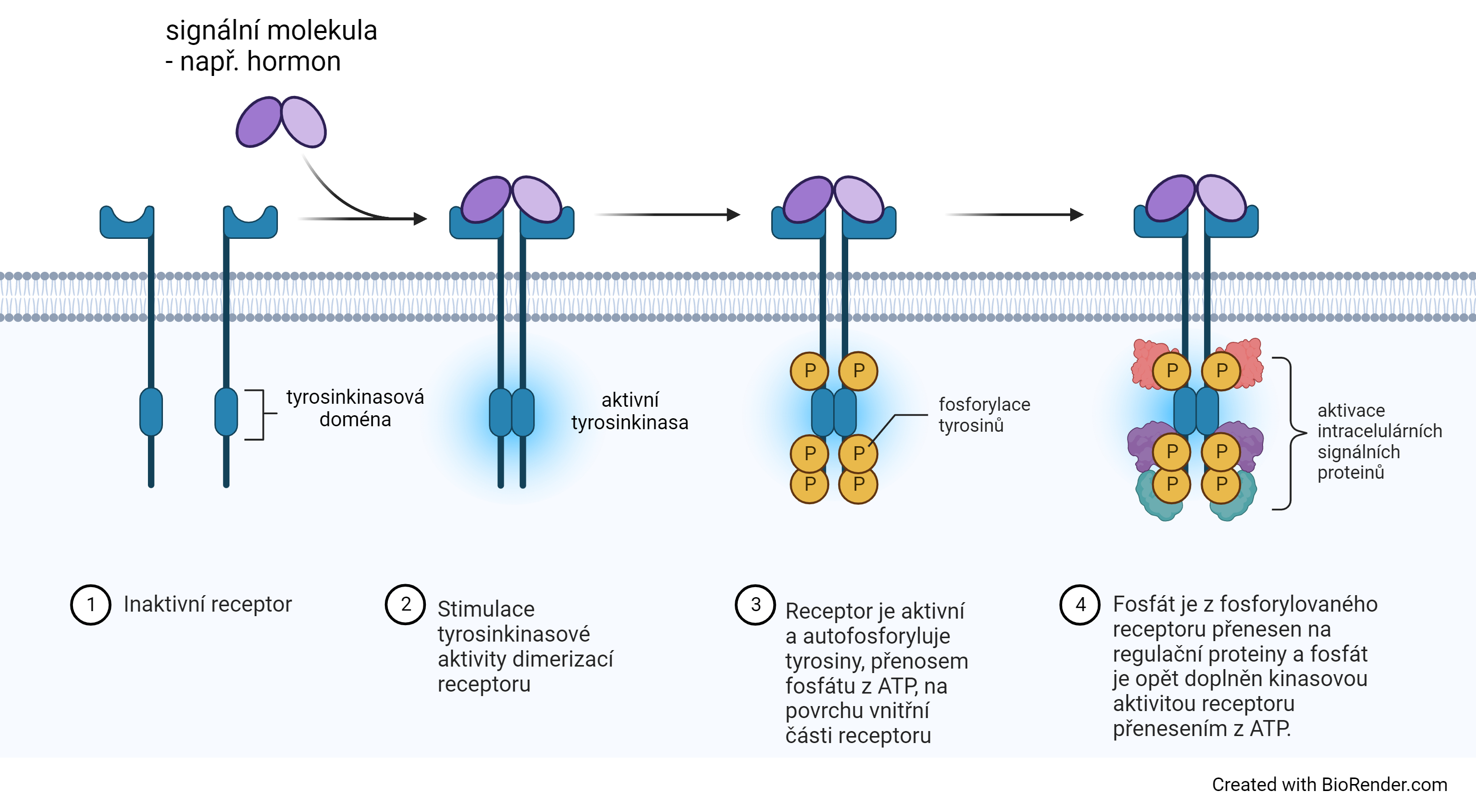 | |
receptoryangl. receptors, bílkoviny, které specificky vážou jiné molekuly (ligandy), přičemž tato  vazba v nich vyvolá mechanismem indukovaného přizpůsobení konformační změnu, která ovlivní jejich chování a v konečném důsledku vyvolá biologickou odpověď buňky. Tak např. vazba v nich vyvolá mechanismem indukovaného přizpůsobení konformační změnu, která ovlivní jejich chování a v konečném důsledku vyvolá biologickou odpověď buňky. Tak např.
| |
receptory 7TMpsáno receptory 7TM, angl. 7 transmembrane receptors nebo seven-transmembrane domain receptors, často se vyskytující jednořetězcové membránové receptory obsahující sedm transmembránových
α-helikálních hydrofobních úseků procházejících lipidovou dvojvrstvou. Tato struktura je typická např. pro receptory interagující s G-proteiny. | |
redoxní systém NAD(P)+, NAD(P)Hangl. redox system NAD(P)+, NAD(P)H, společné označení
NAD+ a NADP+, resp. NADH a NADPH. Tyto koenzymy oxidoreduktas mají několik společných znaků:
| |
reduktasyangl. reductases, triviální název skupiny enzymů ze třídy
oxidoreduktas, u nichž je metabolicky významná redukce určitého substrátu, např.: - enoyl-CoA-reduktasa (EC 1.3.1.39; EC 1.3.1.10, převádějící při biosyntéze mastných kyselin nenasycený enoyl-CoA na nasycený acyl-CoA). | |
regulace transkripceangl. transcription regulation, jeden z nejdůležitějších regulačních mechanismů buněk i celých organismů. Na této úrovni se rozhoduje o syntéze enzymů, potřebných k metabolismu (viz lac-operon), o diferenciaci buněk, o účinnosti hormonální regulace prostřednictvím steroidních hormonů, o syntéze bílkovinných hormonů atd. | |
Reichstein Tadeus | |
rekombinace DNAangl. DNA recombination, buněčný proces, při němž vzniká dceřiná
DNA obsahující segmenty dvou rodičovských DNA. Tento pojem se používá i pro označení procesů, při nichž je in vitro do DNA jednoho druhu vložen úsek DNA jiného druhu tak, aby tato rekombinantní DNA mohla poté být vložena do buňky téhož nebo jiného druhu. Tento postup se v genovém inženýrství využívá pro vložení určitého genu do vektoru, který se nejdříve pomocí restrikční endonukleasy otevře a pak pomocí DNA-ligasy uzavře tak, aby obsahoval nový gen (viz rekombinantní bílkoviny). Za základní studie biochemie nukleových kyselin, zvláště rekombinantní DNA, obdržel roku 1980 P. Berg Nobelovu cenu. | |
rekombinantní DNA | |
renaturaceangl. renaturation, proces, při němž je roztok denaturovaného
biopolymeru zpětně převeden do nativních podmínek (např. z vysoké teploty na teplotu pokojovou, z extrémního pH do pH neutrálního, je z něj odstraněno denaturační činidlo apod.). Přitom může biopolymer zaujmout svou nativní strukturu (vratná denaturace), mnohdy se mu však nativní strukturu nepodařínalézt (denaturace nevratná). | |
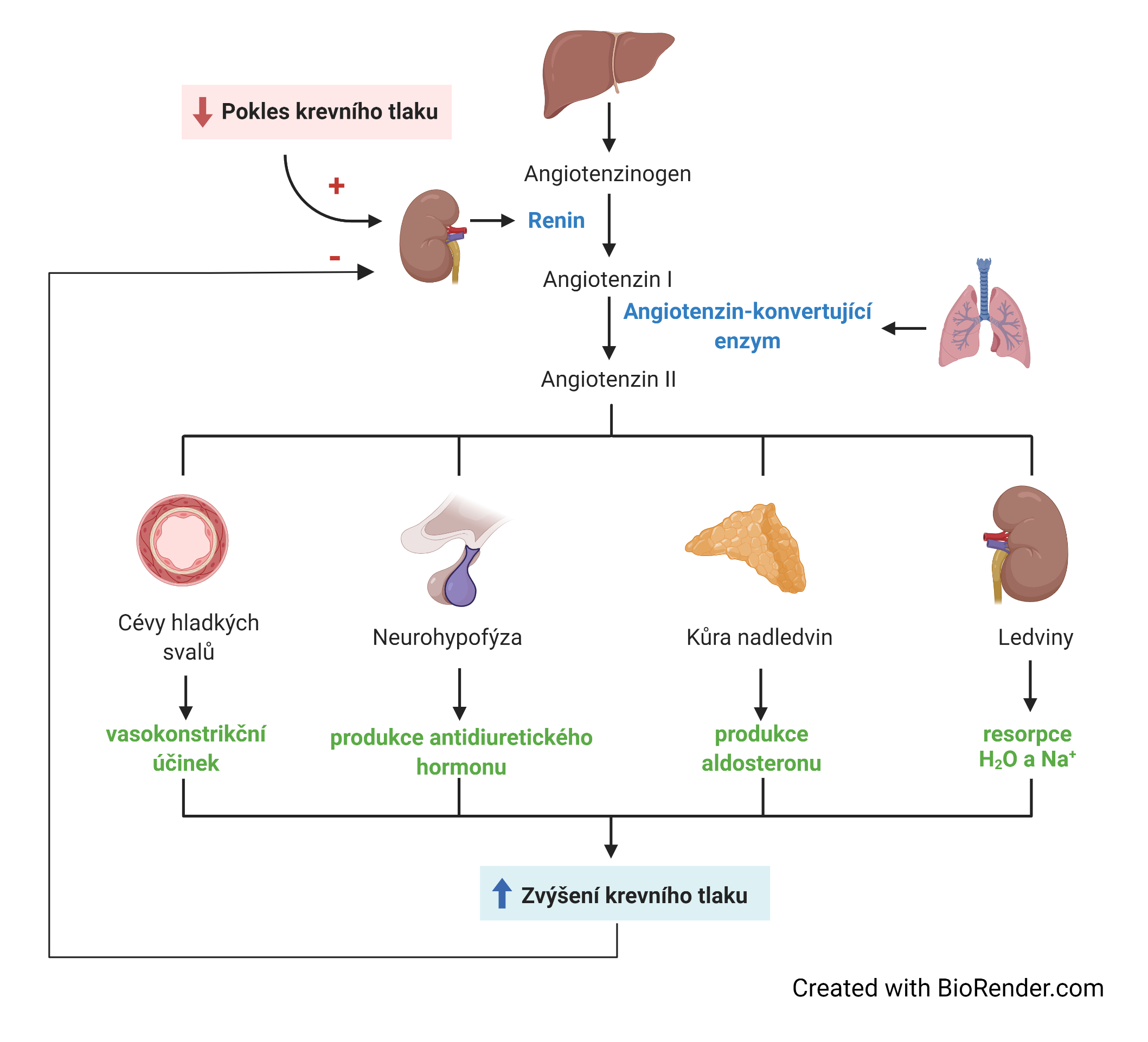
renin-angiotensinový systémtéž systém renin-angiotensin-aldosteron, angl. renin-angiotensin-aldosterone system (RAAS), systém zajišťující homeostázu regulací krevního tlaku, koncentrací solí a množství extracelulární tekutiny. V případě snížení krevního tlaku a/nebo koncentrace sodných iontů v krevní plasmě se začne v ledvinách uvolňovat renin (hormon s proteolytickou aktivitou), který následně z globulinu angiotensinogenu, produkovaného játry, odštěpí dekapeptid angiotensin I. Z něj angiotensin-konvertující enzym, který se nachází především ve stěně plicních cév, odštěpí dipeptid a převede ho na oktapeptid angiotensin II se silnými vasokonstrikčními účinky – vyvolává smršťování cév a zvyšuje tak krevní tlak; navíc v kůře nadledvin stimuluje produkci aldosteronu, steroidního hormonu, který v nefronech ledvin zvyšuje zpětnou resorbci sodných iontů ze vznikající moče, s čímž je vždy spojeno i zadržováním vody. | |
reparace DNAtéž oprava DNA, angl. DNA reparation, soubor buněčných mechanismů, jimiž se mohou odstranit nežádoucí zásahy do struktury
DNA. Lze tak odstranit náhodné chemické modifikace bází (např. dimery pyrimidinových bází vzniklé působením UV-záření), odstranit chybně připojené báze v průběhu replikace nebo spojit přerušené fosfodiesterové vazby v jednom nebo obou vláknech DNA (viz DNA-ligasy). | |
replikace DNAangl. DNA replication, proces, při němž se v souvislosti s 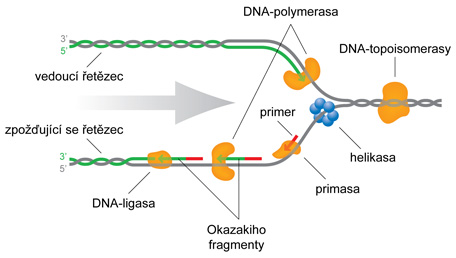 dělením buňky syntetizuje další chemicky totožná molekula
DNA tak, aby obě dceřiné buňky mohly dostat úplnou genetickou informaci. Proces je zahájen v iniciačním bodě přerušením nekovalentních vazeb v dvouřetězcové DNA a vytvořením replikačního očka, kdy se oba řetězce od sebe oddálí. Poté následuje syntéza krátkého úseku RNA (primeru). Vlastní syntézu vlákna DNA řídí enzym DNA-polymerasa, která katalyzuje přenos deoxyribonukleotidů z deoxyribonukleosidtrifosfátů na rostoucí řetězec; protože je zásadně důležité, aby kopie byly dokonale stejné, má DNA-polymerasa schopnost odstranit připojený nukleotid, pokud není připojen komplementárně (viz reparace DNA). Syntéza probíhá od konce 5′ ke konci 3′, proto se jeden řetězec (vedoucí řetězec) syntetizuje kontinuálně a druhý přerušovaně (zpožďující se řetězec, viz Okazakiho fragmenty). Replikace probíhá semikonzervativně; to znamená, že v nově syntetizované molekule je vždy jedno vlákno z původní (dvouřetězcové) molekuly a druhé je nově vytvořené. Za objev mechanismu biosyntézy ribonukleové a deoxyribonukleové kyseliny získali S. Ochoa a A. Kornberg roku 1959 Nobelovu cenu. dělením buňky syntetizuje další chemicky totožná molekula
DNA tak, aby obě dceřiné buňky mohly dostat úplnou genetickou informaci. Proces je zahájen v iniciačním bodě přerušením nekovalentních vazeb v dvouřetězcové DNA a vytvořením replikačního očka, kdy se oba řetězce od sebe oddálí. Poté následuje syntéza krátkého úseku RNA (primeru). Vlastní syntézu vlákna DNA řídí enzym DNA-polymerasa, která katalyzuje přenos deoxyribonukleotidů z deoxyribonukleosidtrifosfátů na rostoucí řetězec; protože je zásadně důležité, aby kopie byly dokonale stejné, má DNA-polymerasa schopnost odstranit připojený nukleotid, pokud není připojen komplementárně (viz reparace DNA). Syntéza probíhá od konce 5′ ke konci 3′, proto se jeden řetězec (vedoucí řetězec) syntetizuje kontinuálně a druhý přerušovaně (zpožďující se řetězec, viz Okazakiho fragmenty). Replikace probíhá semikonzervativně; to znamená, že v nově syntetizované molekule je vždy jedno vlákno z původní (dvouřetězcové) molekuly a druhé je nově vytvořené. Za objev mechanismu biosyntézy ribonukleové a deoxyribonukleové kyseliny získali S. Ochoa a A. Kornberg roku 1959 Nobelovu cenu. | |
replikonangl. replicon, jednotka ve struktuře DNA, která je tvořena iniciačním bodem a tou částí DNA, která je z tohoto místa replikována. Chromosom eukaryot obsahuje asi 1 000 replikonů. | |
represorangl. repressor, v molekulové genetice
bílkovina, která se váže na regulační oblast DNA, zvanou operátorová, a tím inhibuje transkripci jednoho nebo více strukturních genů (viz lac-operon). | |
respirace aerobníangl. aerobic respiration, typ
buněčné respirace, kde konečným akceptorem elektronů (oxidačním činidlem) je molekulový kyslík (srov. anaerobní respirace). | |
respirace anaerobníangl. anaerobic respiration, typ buněčné respirace, kde v dýchacím řetězci není konečným akceptorem elektronů (oxidačním činidlem) molekulový kyslík, ale jiná molekula (částice), přijímaná z extracelulárního prostředí, např. NO3− (denitrifikační bakterie), SO42− (desulfurikační bakterie), CO2 (methanogenní bakterie), fumarát (Escherichia coli), ale také kovy jako Fe3+, Mn4+, Co3+ nebo U6+. Všechna tato oxidační činidla mají nižší standardní redoxní potenciál než O2, a proto je anaerobní respirace energeticky méně výhodná než respirace aerobní. S tímto typem respirace se setkáváme u nefermentujících anaerobních prokaryot; je zde vázána na cytoplasmatickou membránu. | |
respirace buněčnáangl. cell respiration, konečný stupeň katabolismu respirujících chemoorganotrofních organismů, resp. buněk. Chemická energie, dočasně uložená v molekulách redukovaných přenašečů vodíku (v
kofaktorech oxidoreduktas NADH a FADH2), je zde uvolňována jejich oxidací, využita pro generování proton-motivní síly (srov. dýchání) a následně uložena do makroergické sloučeniny, obvykle ATP. | |
retikulum endoplasmatickéz lat. reticulum = síťka, angl. endoplasmatic reticulum nebo endoplasmic reticulum, zkr. ER, složitá organela tvořená systémem biomembrán. Probíhá v něm řada buněčných procesů, především glykosylace proteinů a biosyntéza lipidů. Na povrchu určitých oblastí jeho membrán jsou vázány ribosomy (hrubé nebo drsné ER, angl. rough ER); polypeptidové řetězce, které jsou zde syntetizovány, pronikají do ER, kde jsou posttranslačně modifikovány (viz částice rozpoznávající signál). Jako hladké (angl. smooth) se označuje ER bez ribosomů. Vnitřní prostory ER (nazývají se lumen) obsahují vysokou koncentraci vápenatých iontů; prostřednictvím ovlivňování propustnosti membrány ER pro Ca2+ je regulována jejich koncentrace v cytosolu. | |
retikulum sarkoplasmatickéangl. sarcoplasmatic reticulum nebo sarcoplasmic reticulum, síť membránových tubulů obklopujících myofibrily (aktomyosinová mikrofilamenta) buněk kosterních svalů, specializovaná část
endoplasmatického retikula těchto buněk. Jejich hlavní funkcí je shromažďovat ionty Ca2+ a na příslušný impuls je uvolňovat; zvýšení lokální koncentrace Ca2+ je pak bezprostřední příčinou kontrakce myofibril. | |
retroviryangl. retroviruses, skupina
RNA-virů, které pomocí reverzní transkripce přepisují svůj genom do podoby dvouřetězcové DNA a tu pak začleňují do chromosomu hostitelské buňky. Mezi retroviry nebezpečné pro člověka patří především HIV (angl. human immunodeficiency virus) a některé viry vyvolávající zhoubné bujení (rakovinu). | |
rezonance nukleární magnetickáangl. nucler magnetic resonance, NMR, metoda vycházející ze sledování interakce atomových jader molekul s elektromagnetickým zářením v silném homogenním magnetickém poli. V biochemii se využívá zejména ke studiu prostorového uspořádání biopolymerů, především bílkovin. Její výhodou je možnost studia konformace bílkovin v roztoku, a tím i sledování jejich konformační dynamiky, nevýhodou je skutečnost, že možnost určení prostorového uspořádání bílkoviny je omezeno její molekulovou hmotností (v současné době 20 až 30 kDa). Za příspěvek k rozvoji metody NMR s vysokým rozlišením získal Nobelovu cenu R. R. Ernst v roce 1991, v roce 2002 pak K. Wüthrich za vývoj NMR jako metody umožňující určení trojrozměrné struktury biologických makromolekul v roztoku. | |
rhodopsinangl. rhodopsin, složená transmembránová bílkovina ve světločivných buňkách, tvořená polypeptidovou částí opsinem a prostetickou skupinou retinalem. Absorpce fotonu v něm vyvolává konformační změnu, jejímž následkem je změna membránového potenciálu, následně převedená na optický vjem. | |
riboflavinangl. riboflavin, viz vitamin B2. | |
ribonukleasytéž RNasy, angl. ribonucleases, enzymy hydrolyzující
fosfodiesterové vazby RNA (EC 3.1.13.-; EC 3.1.14.-; EC 3.1.15.-; EC 3.1.16.-; EC 3.1.26.-; EC 3.1.27.-). Trávicí pankreatická RNasa, malá bílkovina (13,7 kDa) s dlouho známou prostorovou strukturou, se stala častým objektem fyzikálně chemických studií. Cytosolové RNasy způsobují, že buněčná RNA (zejména mRNA) má relativně krátký biologický poločas. Za příspěvek k pochopení vztahu mezi chemickou strukturou a katalytickou aktivitou RNasy získali roku 1972 S. Moor a W. H. Stein Nobelovu cenu; ve stejném roce ji dostal také Ch. B. Anfinsen, který na základě studia RNasy objasnil základní vztahy sekvence a prostorového uspořádání proteinů. | |
ribosaangl. ribose, nejdůležitější aldopentosa. Ve furanosové formě je základní součástí 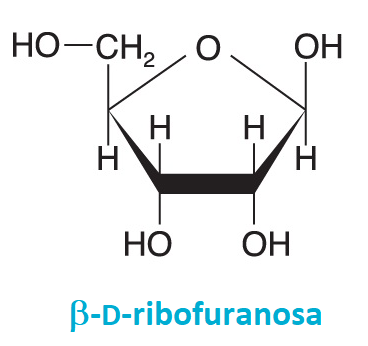 ribonukleosidů a tím i RNA. Fosforylované deriváty ribosy jsou významnými metabolity: ribosa-5-fosfát je meziproduktem pentosového a Calvinova cyklu, 5-fosforibosyl-1-difosfát je meziproduktem biosyntézy purinových a pyrimidinových nukleotidů. 2-Deoxyribosa je součástí cukr-fosfátové páteře řetězce DNA. ribonukleosidů a tím i RNA. Fosforylované deriváty ribosy jsou významnými metabolity: ribosa-5-fosfát je meziproduktem pentosového a Calvinova cyklu, 5-fosforibosyl-1-difosfát je meziproduktem biosyntézy purinových a pyrimidinových nukleotidů. 2-Deoxyribosa je součástí cukr-fosfátové páteře řetězce DNA. | |
ribosomyangl. ribosomes, nejsložitější
nukleoproteinové komplexy buňky; 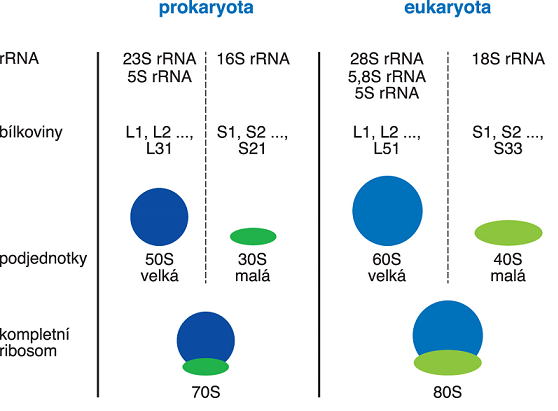 na jejich povrchu dochází k translaci. Obsahují několik typů RNA (viz rRNA) a několik desítek specifických bílkovin. Jsou tvořeny dvěma nestejně velikými nekovalentně spojenými podjednotkami. Nacházejí se buď volné v cytosolu nebo připojené k membráně endoplasmatického retikula. Ribosomy prokaryot, mitochondrií a chloroplastů jsou menší než ribosomy eukaryot (na obr. je velikost, resp. hmotnost částic vyjádřena v jednotkách S - viz sedimentační koeficient). V buňce je jich přítomno velké množství, u bakterie Escherichia coli tvoří až 30 % sušiny. Za objasnění struktury a funkce ribosomů získali v roce 2009 V. Ramakrishnan, T. A. Steitz a A. Yonath Nobelovu cenu. na jejich povrchu dochází k translaci. Obsahují několik typů RNA (viz rRNA) a několik desítek specifických bílkovin. Jsou tvořeny dvěma nestejně velikými nekovalentně spojenými podjednotkami. Nacházejí se buď volné v cytosolu nebo připojené k membráně endoplasmatického retikula. Ribosomy prokaryot, mitochondrií a chloroplastů jsou menší než ribosomy eukaryot (na obr. je velikost, resp. hmotnost částic vyjádřena v jednotkách S - viz sedimentační koeficient). V buňce je jich přítomno velké množství, u bakterie Escherichia coli tvoří až 30 % sušiny. Za objasnění struktury a funkce ribosomů získali v roce 2009 V. Ramakrishnan, T. A. Steitz a A. Yonath Nobelovu cenu. | |
ribozymyangl. ribonucleic enzymes nebo ribozymes, ribonukleové kyseliny s katalytickou aktivitou. Byly popsány především ribozymy působící na RNA (viz sestřih RNA); jistou enzymovou funkci vykazují i rRNA. Existence RNA s enzymovou aktivitou vede k domněnce, že prvotními biopolymery v evoluci mohly být nukleové kyseliny a že tedy proteiny vznikly později. Za objev katalytických vlastností RNA získali roku 1989 S. Altman a T. R. Cech Nobelovu cenu. | |
RNAribonukleová kyselina, angl. ribonucleic acid, lineární polymer ribonukleotidů (A,G,C,U), spojených fosfodiesterovými vazbami. Na rozdíl od DNA je pouze jednořetězcová. Je syntetizována procesem transkripce podle DNA, v níž je uložena informace o pořadí ribonukleotidů ve vznikající RNA (princip komplementarity bází). RNA většinou podléhá dalším úpravám (viz posttranskripční modifikace). Rozlišujeme několik základních typů RNA, které se liší prostorovou strukturou a zejména funkcí:
| |
RNA-interference
zkr. RNAi, angl. RNA interference, mechanismus, jímž jsou v buňkách likvidovány dvouřetězcové molekuly RNA (double-stranded RNA, dsRNA). Předpokládá se, že tento mechanismus vznikl jako obrana buněk proti infekci virovou dsRNA. Při využití v genovém inženýrství může být specifická dsRNA do buňky vpravena mikroinjekcí, elektroporací apod. V současnosti se tato metoda hojně využívá k cílenému útlumu exprese určitých genů (gene knock-out, gene silencing nebo RNA silencing). Využívá se toho, že vložený řetězec RNA, který je komplementární k buněčné mRNA, s ní může vytvořit dsRNA. Tato dvouřetězcová RNA je pak mechanismem RNAi likvidována a nemůže vstoupit do procesu translace.
Nejznámější mechanismus RNAi je následující: 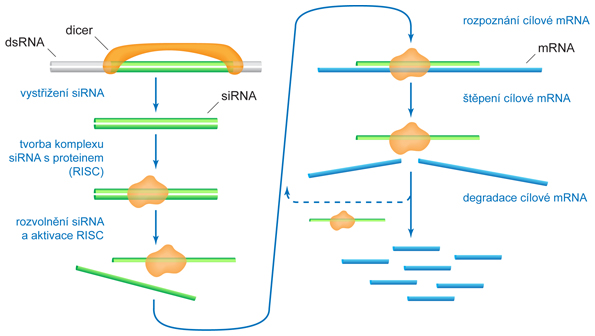 1. dsRNA je v buňce rozštěpena specifickou endonukleasou zvanou DICER. Vznikají tak krátké dvouřetězcové oligonukleotidy obsahující 20 až 25 párů bází, zvané siRNA (short-interfering RNA). 2. siRNA se naváže na specifický proteinový komplex RISC (RNA-induced silencing complex), který řetězce oddělí od sebe a na jeden z nich (antisense RNA) naváže molekulu komplementární mRNA. 3. Takto vzniklý komplex vykazuje endonukleasovou aktivitu a je schopen navázanou mRNA rozštěpit a znemožnit, aby byla použita jako templát pro translaci. Fragmenty mRNA se odpojí a celý proces (od bodu 2) se může opakovat. Tímto způsobem může jedna molekula siRNA pomoci vyhledat a inaktivovat mnoho molekul určité mRNA. 4. Specifická RNA-dependentní-RNA-polymerasa, vyskytující se u všech eukaryot kromě hmyzu a savců, může jednotlivé řetězce siRNA navázané na mRNA využít jako primery a prodloužit dvouřetězcové úseky RNA. Ty se pak stanou substrátem pro DICER (viz bod 1) a cyklus se opakuje. Jedna molekula antisense RNA (nebo dsRNA) tak může způsobit likvidaci velkého počtu molekul mRNA.
Dalším typem antisense RNA vyskytujícím se přirozeně v buňkách je pre-micro RNA. Tyto molekuly jsou syntetizovány podle určitých nekódujících úseků DNA a podílejí se na regulaci exprese určitých genů. Obsahují vlásenkovité struktury, které připomínají dsRNA, a jsou proto štěpeny DICERem za vzniku mikro RNA (miRNA), která se váže na RISC. Mohou zabránit translaci mRNA, která je k jednomu z řetězců komplementární. Za objev RNA-interference (tlumení exprese genů dvouřetězcovou RNA) získali roku 2006 A. Z. Fire a C. C. Mello Nobelovu cenu. | |
RNA-polymerasapřesněji DNA-dependentní-RNA-polymerasa, EC 2.7.7.6, angl. RNA polymerase, enzym ze třídy
transferas katalyzující biosyntézu RNA v procesu transkripce: (NMP)n + NTP → (NMP)n+1 + PPi , kde (NMP)n je prodlužující se oligoribonukeotid a NTP ribonukleosidtrifosfát. RNA-polymerasa je tvořena několika podjednotkami s některými specifickými funkcemi; např. σ-podjednotka u prokaryot zajišťuje vazbu enzymu na promotor, a tím iniciaci transkripce. | |
Robinson Robert | |
Rodbell Martin
| |

Rose Irwinamerický biochemik ( | |

rostliny C3, C4 a CAMangl. C3, C4 a CAM plants, rozlišení rostlin podle způsobu fixace
CO2 při fotosyntéze. U rostlin C3 reaguje CO2 s ribulosa-1,5-bisfosfátem za katalýzy enzymem RUBISCO a prvním detekovatelným produktem je tříuhlíkatý (proto C3) meziprodukt glukogenese 3-fosfoglycerát. Do této skupiny patří většina rostlin mírného pásma. Nevýhodou tohoto způsobu fixace CO2 je to, že v důsledku nízkého parciálního tlaku CO2 v atmosféře a vysokého parciálního tlaku O2 zde probíhá poměrně intenzivní fotorespirace. U rostlin C4 je primárním akceptorem CO2 fosfoenolpyruvát a vzniká čtyřuhlíkatý oxalacetát (viz cyklus C4-rostlin); mezi C4-rostliny patří mnohé tropické rostliny, z našich pak zejména kukuřice. Rostliny CAM (Crassulacean Acid Metabolism – metabolismus kyselin u tučnolistých) v noci fixují CO2 podobným způsobem jako C4-rostliny, uchovávají malát ve vakuolách a ve dne, kdy je dostatek světla a tedy i ATP a NADPH, realizují Calvinův cyklus; je tedy u nich časově oddělena fixace CO2 od jeho využití. Tyto rostliny, např. kaktusovité nebo agávovité, jsou adaptovány na horké a suché podnebí tím, že během dne mohou mít uzavřené průduchy a brání se tak vysychání. | |
Rothman James Edward
| |
 americký buněčný biolog (nar. 1950), Nobelova cena (za fyziologii a lékařství, 2013) za studii regulace
americký buněčný biolog (nar. 1950), Nobelova cena (za fyziologii a lékařství, 2013) za studii regulace rovnice Goldmanova
též Goldmanova–Hodgkinova–Katzova rovnice, angl. Goldman equation, vztah umožňující vypočítat membránový potenciál (Δφ) pomocí intracelulárních (i) a extracelulárních (e) koncentrací nejdůležitějších iontů (Na+, K+, Cl−) a pomocí koeficientůbpropustností P těchto iontů membránou:
\( \Delta \phi= \frac{RT}{F} ln \frac{ P_{Na} [ Na^{+}]_e +P_{K} [ K^{+}]_e + P_{Cl} [ Cl^{-} ]_i }{P_{Na} [ Na^{+}]_i +P_{K} [ K^{+}]_i + P_{Cl} [ Cl^{-} ]_e} \) Vzhledem k tomu, že koncentrace iontů na obou stranách membrány se příliš nemění, jsou prudké změny membránového potenciálu, charakteristické např. pro vedení nervového vzruchu, způsobeny náhlými změnami pasivní propustnosti iontů (viz pasivní transport, iontové kanály). | |
rovnice Hillovaangl. Hill equation, vztah mezi počáteční rychlostí enzymové reakce v0 u allosterických enzymů a koncentrací substrátu [S]: \( v_{0} = \frac{ V_{lim} . [S] ^{n} }{ K^{ `} + [S]^{n} } \) kde vlim je limitní rychlost (viz rovnice Michaelise a Mentenové), a K′ je konstanta, jen formálně připomínající Michaelisovu konstantu. Parametr n (často též označovaný α) se nazývá koeficient sigmoidity nebo též Hillův; čím vyšší má hodnotu, tím více se závislost v0 na [S] liší od hyperboly a má výraznější tvar sigmoidy. Hodnota n bývá často rovna počtu enzymově aktivních podjednotek v oligomerním enzymu, její hodnota roste při allosterické inhibici a klesá při allosterické aktivaci. Formálně podobným vztahem se řídí i závislost rozsahu vazby nízkomolekulárního ligandu na oligomerní vazebnou bílkovinu s kooperujícími podjednotkami (např. vazby kyslíku na hemoglobin). Závislost počáteční reakční rychlosti na koncentraci substrátu u allosterických enzymů (čísla udávají hodnoty koeficientu sigmoidity n) 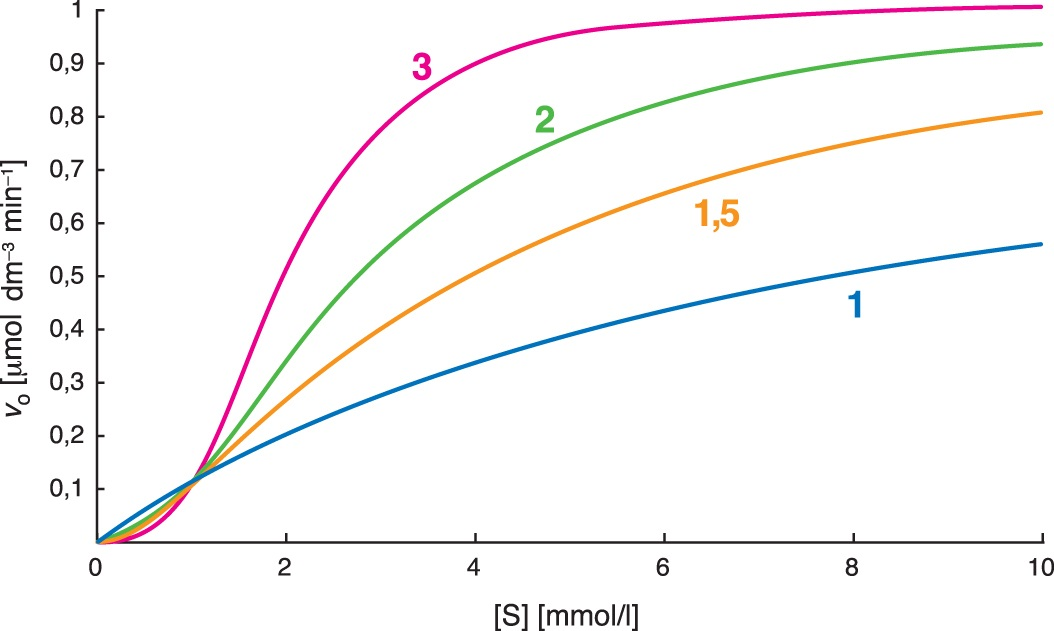 | |
rRNAribosomální
RNA, angl. ribosomal RNA, RNA podílející se spolu se specifickými bílkovinami na výstavbě ribosomu. Je nejhojnějším typem RNA (až 80 hmotn. % všech RNA). Jedná se o několik druhů molekul lišících se velikostí a zastoupením jednotlivých nukleotidů. U eukaryot je syntetizována v jadérku. Vytváří některé specifické sekundární struktury (vlásenky nebo dihelixy podobné DNA). Spolu s ribosomálními proteiny (viz peptidyltransferasa) se podílí na katalýze translace; lze ji tedy považovat za ribozym. | |
RUBISCOribulosa-1,5-bisfosfátkarboxylasa/oxygenasa, angl. též ribulose-1,5-bisphosphate carboxylase/oxygenase, EC 4.1.1.39, enzym ze třídy lyas, který v temné fázi fotosyntézy katalyzuje připojení CO2 k molekule ribulosa-1,5-bisfosfátu a následné rozštěpení vzniklého meziproduktu na dvě molekuly 3-fosfoglycerátu. Kromě této karboxylační reakce (fixace CO2) katalyzuje oxygenační reakci (viz fotorespirace), při níž se na molekulu stejného akceptoru váže molekulový kyslík a kromě 3-fosfoglycerátu vzniká fosfoglykolát. Z něho se sice v rostlinách syntetizuje serin a glycin, většina se však katabolicky odbourává a z hlediska celkové bilance je to ztrátová reakce. 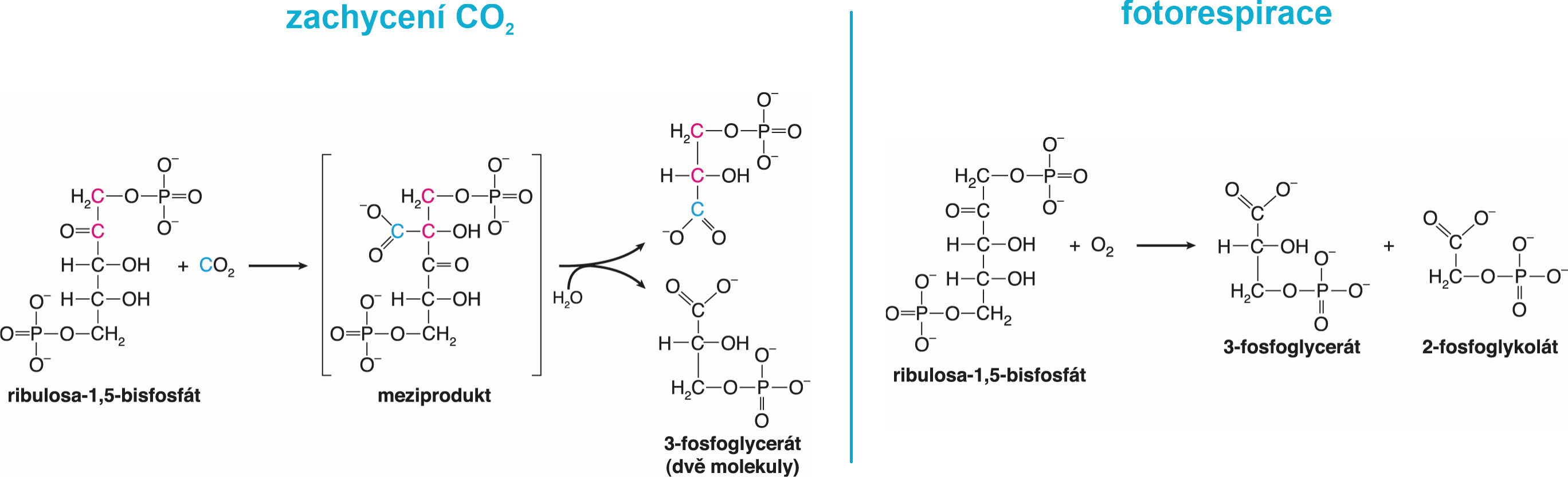 | |
Ružička Leopold
| |
 chorvatský (rakousko
chorvatský (rakouskorychlost limitnívlim, nebo maximální vmax , angl. limiting rate nebo velocity, jeden ze základních parametrů rovnice Michaelise a Mentenové (viz enzymy – kinetika), teoreticky nejvyšší počáteční reakční rychlost, jíž může reakce při dané koncentraci enzymu za určitých podmínek (pH, teploty atd.) dosáhnout. Z různých důvodů (inhibice substrátem, omezená rozpustnost substrátů atd.) nemusí být možné této rychlosti experimentálně dosáhnout. | |
rychlost reakční počátečníangl. initial reaction rate nebo initial reaction velocity, pojem užívaný v enzymové kinetice pro rychlost reakce v čase
t = 0, tedy pro rychlost, kterou lze změřit po přidání enzymu do roztoku substrátů. Koncentrace produktů je v tomto okamžiku nulová a reakce tedy probíhá pouze jedním směrem. | |