Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
P |
|---|
Pääbo Svante
| |
 švédský evoluční genetik, nar. 1955, Nobelova cena (za fyziologii a lékařství, 2022) za objevy, které se týkají
švédský evoluční genetik, nar. 1955, Nobelova cena (za fyziologii a lékařství, 2022) za objevy, které se týkají papainangl. papain, proteolytický enzym obsažený v nezralých plodech a latexu papajovníku (Carica papaya), EC 3.4.22.2. Katalytickou skupinou v aktivním centru je skupina SH v cysteinu. Jeho prostorová struktura je mimořádně stabilní, denaturuje při vyšší teplotě než živočišné enzymy, odolává denaturaci osmimolární močovinou a 70% ethanolem. Hydrolyticky štěpí peptidy, amidy i estery; v peptidovém řetězci nejlépe štěpí druhou peptidovou vazbu následující směrem k C-konci za zbytkem fenylalaninu. Používá se k tenderizování masa, jako součást tablet napomáhajících trávení a v kožním lékařství při léčení ekzémů a nekróz. | |
parathormontéž parathyrin, angl. parathyroid hormone nebo parathormone, zkr. PTH, peptidový hormon (84 aminokyselin) vylučovaný příštítnými tělísky. Řídí metabolismus vápníku a fosfátu. Ovlivňuje buňky, které degradují kost (osteoklasty). Mobilizace těchto buněk zvyšuje koncentraci vápenatých iontů v krvi a následně snižuje koncentraci fosfátových iontů. Nedostatek parathormonu vyvolává křeče, jeho úplný deficit smrt. | |
párování bázíangl. base pairing, schopnost
bází nukleových kyselin vytvářet pomocí vodíkových můstků komplementární páry, tedy guanin-cytosin, adenin-thymin a adenin-uracil (viz komplementarita bází). Toto specifické vzájemné rozpoznávání je základem přenosu genetické informace, neboť umožňuje jak syntézu totožné molekuly DNA při dělení buněk (viz replikace), tak syntézu peptidového řetězce bílkoviny podle informace zapsané v genu (viz transkripce a translace). Pojem pár bází, zkr. bp, se (poněkud kuriózně) používá jako jednotka délky dihelixu DNA; např. údaj úsek 160 párů bázíznamená, že tento úsek obsahuje 160 komplementárních dvojic deoxyribonukleotidů. | |
Patapoutian Ardem
| |
 americký molekulární biolog a neurovědec (nar. 1967), Nobelova cena (za fyziologii a lékařství, 2021) za objev
americký molekulární biolog a neurovědec (nar. 1967), Nobelova cena (za fyziologii a lékařství, 2021) za objev Pauling Linus Carl americký chemik (1901–1994), Nobelova cena (chemie, 1954) za výzkum povahy chemické vazby a aplikaci získaných poznatků pro určení struktury komplexních sloučenin. Narodil se v Portlandu (Oregon), studoval chemické inženýrství, později se plně věnoval teoretické fyzice. Šíře jeho vědeckých zájmů je mimořádná: pomocí rentgenové krystalografie studoval struktury aminokyselin a peptidů, zabýval se aplikací kvantové mechaniky při popisu chemické vazby, při studiu struktury proteinů popsal základní periodické motivy peptidového řetězce, studoval strukturu protilátek a povahu serologické reakce, jakož i strukturu abnormálního hemoglobinu ve vztahu k hemolytickým anémiím. Veřejnosti je znám jako propagátor použití masivních dávek vitaminu C jako „univerzálního“ prostředku pro ochranu zdraví. V roce 1962 získal Nobelovu cenu míru za úsilí o zákaz zkoušek jaderných zbraní; je dosud jediným člověkem, který dvakrát získal nedělenou Nobelovu cenu. americký chemik (1901–1994), Nobelova cena (chemie, 1954) za výzkum povahy chemické vazby a aplikaci získaných poznatků pro určení struktury komplexních sloučenin. Narodil se v Portlandu (Oregon), studoval chemické inženýrství, později se plně věnoval teoretické fyzice. Šíře jeho vědeckých zájmů je mimořádná: pomocí rentgenové krystalografie studoval struktury aminokyselin a peptidů, zabýval se aplikací kvantové mechaniky při popisu chemické vazby, při studiu struktury proteinů popsal základní periodické motivy peptidového řetězce, studoval strukturu protilátek a povahu serologické reakce, jakož i strukturu abnormálního hemoglobinu ve vztahu k hemolytickým anémiím. Veřejnosti je znám jako propagátor použití masivních dávek vitaminu C jako „univerzálního“ prostředku pro ochranu zdraví. V roce 1962 získal Nobelovu cenu míru za úsilí o zákaz zkoušek jaderných zbraní; je dosud jediným člověkem, který dvakrát získal nedělenou Nobelovu cenu. | |
pektinyangl. pectins, polysacharidy obsahující jako monomerní jednotky především galakturonovou kyselinu a její estery, ale také rhamnosu a řadu dalších monosacharidů. Jsou rozšířeny v rostlinné říši; jsou vázány na polysacharidy buněčných stěn a tvoří buněčný tmel (střední lamelu). Získávají se z řepných řízků, z jablek a citrusových plodů. Díky přítomnosti silně hydrofilních karboxylových skupin mají pektiny velkou schopnost vázat vodu a za určitých podmínek (pH, koncentrace Ca2+) vytvářejí gely. Toho se využívá v potravinářství (při výrobě marmelád), ve farmaceutickém průmyslu a v kosmetice. | |
pentosyangl. pentoses, monosacharidy obsahující řetězec pěti atomů uhlíku. Aldopentosy se vyskytují v relativně stabilní cyklické (furanosové) podobě. Nejvýznamnější pentosy jsou aldopentosy
ribosa a 2-deoxyribosa (tvoří základ nukleosidů), arabinosa a xylosa. V metabolismu sacharidů hrají významnou roli fosforečné estery ketopentos, ribulosa-5-fosfát a xylulosa-5-fosfát. Ribulosa-1,5-bisfosfát je akceptorem oxidu uhličitého v temné fázi fotosyntézy. Pentosy metabolicky vznikají oxidačním zkrácením hexos v pentosovém cyklu. | |
pepsinangl. pepsin, proteolytický
enzym (EC 3.4.23.1, EC 3.4.23.2, EC 3.4.23.3) obsažený v žaludku všech obratlovců (kromě některých ryb, např. kapra). Má nejvyšší aktivitu při nízkém pH, odpovídajícím podmínkám v žaludku (1,5 až 3,5); nad pH 6 je inaktivován denaturací. Nemá velkou specifitu, nejlépe štěpí peptidové vazby na karboxylové straně aromatických aminokyselin (Tyr, Phe). Produktem působení pepsinu na bílkoviny je pepton, směs peptidů obsahujících obvykle 3 až 30 aminokyselinových zbytků. Pepsin je syntetizován buňkami žaludeční sliznice ve formě proenzymu pepsinogenu, který je v kyselém prostředí autokatalyticky aktivován. | |
peptidasyangl. peptidases, enzymy ze třídy
hydrolas (EC 3.4.-.-), štěpící peptidovou vazbu. Do této rozsáhlé skupiny řadíme aminopeptidasy, karboxypeptidasy, dipeptidasy, proteinasy a mnohé další trávicí a regulační enzymy. | |
peptidyangl. peptides, kondenzační produkty
aminokyselin, které jsou spojeny peptidovou vazbou. Dělíme je podle řady kritérií:
| |
peptidyltransferasaangl. peptidyltransferase, enzym katalyzující vznik
peptidové vazby při translaci, součást velké podjednotky ribosomů, EC 2.3.2.12. Přenáší peptidylový zbytek, vázaný prostřednictvím tRNA v P-místě ribosomu, na volnou aminoskupinu aminoacyl-tRNA, vázanou v A-místě. | |
permeasyangl. permeases, integrální transmembránové bílkoviny umožňující pasivní specifický přenos malých částic membránou. Do této skupiny bílkovin patří jak iontové kanály, tak transportní systémy pro nenabité nízkomolekulární látky (např. pro glukosu). | |
peroxidasyangl. peroxidases, enzymy ze třídy
oxidoreduktas (EC 1.11.1.-), které pomocí peroxidu vodíku (jako oxidačního činidla) oxidují různé substráty: redukovaný substrát + H2O2 → oxidovaný substrát + 2 H2O. Jsou součástí ochrany organismů proti oxidačnímu stresu, neboť odstraňují peroxid vodíku (viz reaktivní kyslíkové částice). Jejich prostetickou skupinou je většinou hem. Nespecifická křenová peroxidasa (EC 1.11.1.7) se užívá při metodách ELISA k značení protilátek nebo antigenů a v mnoha enzymových stanoveních, při nichž vzniká H2O2. | |
peroxisomangl. peroxisome, organela, v níž jsou lokalizovány mnohé reakce, při nichž vzniká peroxid vodíku (viz oxidasy, např. acyl-CoA-oxidasa), který je zde rychle rozkládán katalasou. V rostlinných peroxisomech probíhá fotorespirace. V podobných organelách rostlinných buněk (viz glyoxysomech) je lokalizován glyoxylátový cyklus. | |
Perutz Max Ferdinand
| |

pigmenty fotosyntetickéangl. photosynthetic pigments, barviva podílející se na zachycení a zužitkování světla při
fotosyntéze. Jsou to především chlorofyly a a b, jakož i různé karotenoidy a fykobiliny. | |
pigmenty respiračnítéž dýchací barviva, angl. respiratory pigments, bílkoviny, které u živočichů zajišťují přísun kyslíku do tkáňových buněk. U obratlovců tuto funkci plní hemoglobin, uzavřený v červených krvinkách, u bezobratlých jiné, obvykle barevné proteinové struktury. | |
pinocytózaangl. pinocytosis, z řeckého
pinein = pít, endocytický proces, při němž je do buňky vnesen roztok s rozpuštěnými látkami, srov. fagocytóza a endocytóza. | |
plasma krevníangl. blood plasma, kapalná složka krve, krev zbavená krevních tělísek (červených krvinek, bílých krvinek a destiček). Tvoří asi
60 % objemu krve. Obsahuje asi 90 % vody, 7 až 8 % bílkovin, 1 % solí, glukosu (viz glykemie) a další nízkomolekulární látky (kyseliny, barviva atd.). | |
plasmalogenyangl. plasmalogens, fosfolipidy strukturně podobné fosfatidylcholinu | |
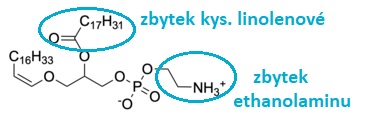 nebo fosfatidylethanolaminu;
nebo fosfatidylethanolaminu; plasmidyangl. plasmids, v
prokaryotech menší úseky DNA, které nejsou součástí bakteriálního chromosomu (hlavního podílu DNA) a replikují se nezávisle na něm (nazývají se také extrachromosomální DNA). Plasmidy tvoří až 5 % prokaryotní DNA, vyskytují se v buňce často v několika (někdy mnoha) kopiích a jsou obvykle cyklické. Jsou v nich uloženy informace o méně závažných a proměnných vlastnostech prokaryotní buňky; obsahují např. geny zajišťující resistenci proti antibiotikům nebo některé specifické metabolické procesy. Mohou se vyměňovat mezi neporušenými buňkami. Upravené plasmidy se používají v mnoha genových manipulacích jako vektory. | |
plasmin | |
plastochinonangl. plastoquinone, isoprenoidní koenzym, součást membrány thylakoidů, obdoba ubichinonu (viz koenzym Q). Účastní se přenosu vodíkových atomů ve světlé fázi fotosyntézy. 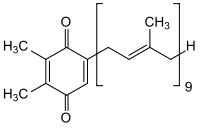 | |
pletivo | |
pohyb flip-flopangl. flip-flop mobility, též příčná difuze (transverse diffusion), pohyb molekul polárních lipidů mezi dvěma vrstvami tvořícími biologické (zejména buněčné) membrány. Vzhledem k energetické náročnosti průniku polární hlavice je tento pohyb zprostředkován membránovými enzymy (flipasami). Tento pohyb lipidovou dvojvrstvou je spojen s "překlopením" molekuly polárního lipidu. | |
polarita molekulyangl. molecular polarity, hodnota dipólového momentu molekuly. Podle tohoto kritéria dělíme molekuly na polární a nepolární; polární molekuly bývají dobře rozpustné v polárních rozpouštědlech, zejména ve vodě, a naopak nepolární molekuly v nepolárních, zejména organických rozpouštědlech (diethyletheru, cyklohexanu apod.). Polární však mohou být i jednotlivé funkční skupiny; pak může být molekula jako celek nepolární, ale přitom ve vodě dobře rozpustná. Za příklad může sloužit hydrochinon (p-dihydroxybenzen), jehož symetrická nepolární molekula obsahuje dvě hydrofilní hydroxylové skupiny, které umožňují jeho relativně velkou rozpustnost ve vodě (5,9 g na 100 ml při 15 °C). Proto je pro vysvětlení interakce molekuly s vodou důležitější pojem hydrofilita než polarita. | |
poloměr van der Waalsůvangl. van der Waals radius, efektivní poloměr atomu, který udává, na jakou nejmenší vzdálenost se k němu může přiblížit jiný atom. Znalost těchto vzdáleností podmiňuje studium těsného sbalení
biopolymerů do nativních struktur. | |
polymorfismus bílkovinangl. protein polymorphism, mnohočetné formy bílkoviny vykazující stejnou funkci a pocházející z různých jedinců téhož druhu nebo z různých orgánů, tkání nebo organel téhož jedince; setkáváme se však i s případy, kdy se polymorfní bílkoviny nacházejí v jednom kompartmentu, např. v krevní plasmě (srov. isoenzymy). Jsou si velmi podobné prostorovým uspořádáním, mají však mírně odlišnou
primární strukturu; jsou tedy kódovány různými geny (srov. alely). | |
polynukleotidyangl. polynucleotides, lineární sekvence více než deseti
nukleotidů spojených fosfodiesterovými vazbami 3′→5′, obvykle synonymum pro nukleové kyseliny. Ve výzkumu se uplatňují různé syntetické polynukleotidy, např. polyA nebo poly(AT). | |
polypeptidyangl. polypeptides, lineární polymery tvořené zbytky
aminokyselin, které jsou spojeny peptidovou vazbou (viz peptidy). Obvykle do této skupiny řadíme peptidy s větším počtem aminokyselinových zbytků než 10 (viz oligopeptidy) a menším než 100 (viz bílkoviny); lze tedy říci, že polypeptidy jsou malé bílkoviny nebo velké oligopeptidy. Zajímavou skupinou polypeptidů jsou syntetické homopolymery aminokyselin (např. poly-L-lysin, poly-L-glutamát), které mohou vysoce převyšovat horní hranici tohoto rozmezí. Nejdůležitější skupinou přirozených polypeptidů jsou polypeptidové hormony (např. insulin, glukagon, hormony adenohypofýzy). | |
polyribosomytéž polysomy, angl. polyribosoms, komplexní struktury tvořené mRNA a několika ribosomy, 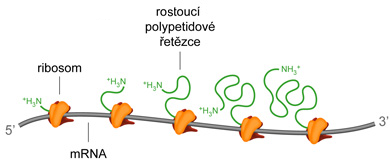 na nich současně probíhá translace. U prokaryot se polyribosomy formují již v průběhu transkripce. na nich současně probíhá translace. U prokaryot se polyribosomy formují již v průběhu transkripce. | |
polysacharidytéž glykany, angl. polysaccharides, sacharidy tvořené více než 10 (obvykle však stovkami až tisíci) monosacharidových jednotek, které jsou spojeny glykosidovými vazbami. Řetězce mohou být větvené nebo nevětvené (lineární). Prostorové uspořádání polysacharidových řetězců může být lineární, helikální nebo globulární. Homoglykany jsou tvořeny jediným druhem monosacharidu, heteroglykany jich obsahují více typů. Polyuronidy obsahují zbytky alduronových kyselin. Nejběžnějšími složkami polysacharidů jsou hexosy D-glukosa (homoglykany celulosa, škroby, glykogen), D-fruktosa (inulin), D-galaktosa (agar-agar), D-mannosa (mannany), D-glukuronová kyselina (hyaluronová kyselina), D-galakturonová kyselina a D-rhamnosa (pektiny), a aminocukr D-glukosamin (chitin, hyaluronová kyselina). Polysacharidy mají především funkci zásobní (škroby, glykogen) a strukturní (např. celulosa, hemicelulosy, chitin, hyaluronová kyselina). | |
pool metabolituvysl. půl, též hotovost nebo fond, angl. pool, hladina důležitých metabolitů v buňce, udržovaná homeostasou na konstantní úrovni. Název pool(angl. bazén) velmi dobře vystihuje skutečnost, že do této hotovosti trvale přitéká a z ní odtéká daný metabolit, ale jeho koncentrace (hladina) zůstává stálá (stav dynamické rovnováhy). Intracelulární stacionární koncentrace důležitých metabolitů bývá řádu mmol.dm-3, jejich obrat (rychlost biosyntézy a odbourávání) však značný. Nejdůležitější buněčné poolytvoří jednotlivé aminokyseliny, acetyl-CoA, ATP, glukosa-6-fosfát, v mitochondrii také meziprodukty citrátového cyklu (viz dráhy anaplerotické). | |
porfyrinyangl. porphyrins, sloučeniny strukturně odvozené od cyklického rovinného 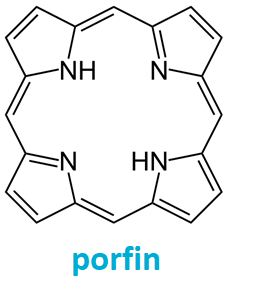 konjugovaného tetrapyrrolu porfinu. Biologicky funkční deriváty mají na kruhové struktuře různé krátké substituenty a uprostřed kruhu koordinován ion kovu:
hem obsahuje železo (Fe2+ v hemoglobinu a myoglobinu, Fe3+ např. v cytochromech, peroxidase a katalase), chlorofyly obsahují hořčík Mg2+ a kobalamin kobalt Co3+. konjugovaného tetrapyrrolu porfinu. Biologicky funkční deriváty mají na kruhové struktuře různé krátké substituenty a uprostřed kruhu koordinován ion kovu:
hem obsahuje železo (Fe2+ v hemoglobinu a myoglobinu, Fe3+ např. v cytochromech, peroxidase a katalase), chlorofyly obsahují hořčík Mg2+ a kobalamin kobalt Co3+. | |
Porter Rodney Robert
| |

póry iontovéangl. ion pores, viz iontové kanály. Někdy se terminologicky odlišují málo specifické iontové póry od mnohem specifičtějších iontových kanálů. | |
póry trvaléangl. permanently open pores, otvory v
biologických membránách, které umožňují snadný přechod látek mezi některými kompartmenty eukaryotních buněk. Díky trvalým pórům je pro molekuly propustná jaderná membrána, jakož i vnější membrána mitochondrie. Otvory v membráně jsou tvořeny specifickými oligomerními bílkovinami, zvanými poriny. | |
posel druhýangl. second messenger, nízkomolekulární intracelulární látka, která se tvoří jako odpověď na
signál, např. na vazbu hormonu ( prvního posla) na receptor. Typickými představiteli této skupiny látek jsou cAMP, cGMP, inositol-1,4,5-trisfosfát a 1,2-diacylglycerol, ale také Ca2+ uvolňovaný do cytoplasmy z endoplasmatického retikula. Druhý posel bývá allosterickým aktivátorem nebo inhibitorem intracelulárních enzymů. | |
potenciál akčníangl. action potential, dočasná prudká změna
membránového potenciálu nervové buňky (z hodnoty klidového potenciálu přibližně −60 mV na hodnotu +30 mV); změna se pohybuje podél axonu nervové buňky a je podstatou vedení nervového vzruchu. Tato změna je vyvolána změnou pasivní propustnosti cytoplasmatické membrány pro sodné a draselné ionty (viz Goldmanova rovnice). | |
potenciál membránovýangl. membrane potential, rozdíl elektrického potenciálu mezi dvěma stranami
biologické membrány (Δφ). Je vyvolán nerovnoměrným rozdělením volných kationtů a aniontů na obou stranách membrány (viz proton-motivní síla) a také přítomností nabitých skupin, které jsou součástí membrány (fosfatidy, sialová kyselina). Membránový potenciál se obvykle pohybuje v jednotkách až desítkách milivoltů, membránový potenciál vnitřní membrány respirující mitochondrie dosahuje hodnot 150 mV. Je-li vnější prostor kompartmentu nabit záporně a vnitřní kladně, považuje se definičně potenciál membrány za kladný (Δφ > 0). Za výzkum elektrických jevů spojených s biologickými membránami získali v roce 1991 E. Neher a B. Sakmann Nobelovu cenu. | |
práce osmotickáangl. osmotic work, energie, kterou musí buňka nebo organismus vynakládat pro udržení koncentračních nerovnováh mezi jednotlivými
kompartmenty a samozřejmě mezi buňkou a extracelulárním prostorem (srov. aktivní transport). Mnohé buňky (erythrocyty, nervové buňky) využívají většinu ATP získaného katabolickými procesy právě na osmotickou práci. | |
práce regulačníangl. regulatory work, energie, kterou musí organismus vynakládat na udržení své vlastní vysoké uspořádanosti, resp. nízké entropie. Typickými procesy vyžadujícími regulační práci je syntéza
hormonů a dalších regulačních látek (např. druhých poslů) nebo energie vynaložená na replikaci, transkripci i translaci. | |
precipitacetéž srážení, angl. precipitation, v biochemii jedna ze základních metod izolace biomakromolekul. Do roztoku obsahujícího požadovanou makromolekulu se přidá určité množství precipitačního činidla (síranu amonného, ethanolu, acetonu apod.), čímž se makromolekulární sloučenina vysráží, nemělo by však dojít k její denaturaci. Může se proto následně znovu rozpustit a použít ve své nativní, biologicky aktivní podobě. | |
pregnenolonangl. pregnenolone, prekursor steroidních hormonů progesteronu, estrogenu, testosteronu, dehydroepiandrosteronu, kortizolu a aldosteronu. Jeho syntéza z cholesterolu je katalyzována oxygenasou ze skupiny cytochrom-P450. | |
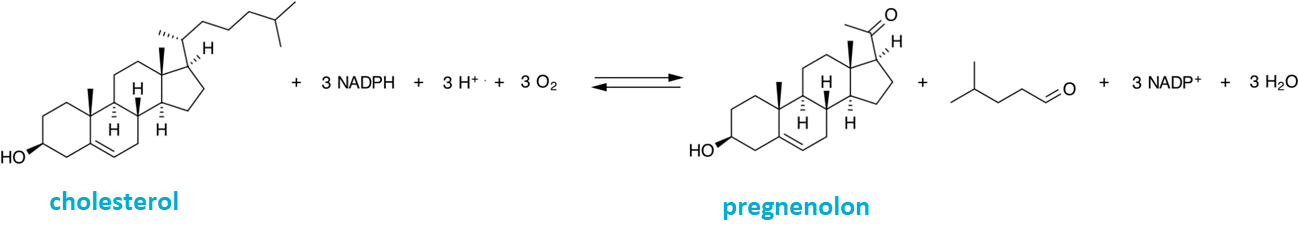
prekursorangl. precursor, sloučenina stojící na počátku biosyntetické dráhy určitého
metabolitu. Tak prekursorem glukosy v játrech je laktát, prekursorem aminokyseliny prolinu meziprodukt citrátového cyklu 2-oxoglutarát atd. Vzhledem k propojenosti metabolických drah mohou mít některé metabolity více prekursorů. Někdy je stanovení skutečného počátečníhoprekursoru věcí osobního pohledu: je prekursorem prolinu L-glutamát, 2-oxoglutarát nebo dokonce glukosa? Jiný příklad: prekursorem všech isoprenoidů je isopentenyldifosfát, jeho prekursorem je však acetyl-CoA. | |
Prigogine Ilya
| |

primerangl. primer, krátký úsek jednořetězcové
DNA nebo RNA, který se na principu párování bází váže k templátovému řetězci DNA; obsahuje volnou skupinu 3′-OH, na níž může DNA-polymerasa navázat další deoxyribonukleotid a zahájit tak vlastní polymerační reakci. Při iniciaci replikace in vivo vytváří primer RNA-polymerasa (též primasa). Syntetické primery tvořené krátkou sekvencí jednořetězcové DNA se užívají při polymerasové řetězové reakci a při sekvenování DNA Sangerovou dideoxy-metodou. | |
prionyangl. prions, bílkoviny vyvolávající choroby (bez účasti
virů nebo bakterií). Tyto bílkoviny jsou přítomné i ve zdravé tkáni, mohou výrazným způsobem změnit konformaci (obvykle z α-helikální na β-konformaci, viz bílkoviny – strukturní motivy) a stát se iniciátorem degenerativního procesu. Jsou příčinou některých onemocnění nervové tkáně (Creuzfeldtovy–Jacobovy nemoci, nemoci šílených krav a dalších). Řada otázek spojená s jejich šířením a působením dosud není vysvětlena. Některé experimentální nálezy z posledního období naznačují, že priony se podílejí na uchovávání paměťové informace v mozku. Za objev prionů ("nového biologického principu infekce") obdržel S .B. Prusiner roku 1997 Nobelovu cenu. | |
proces endergonickýangl. endergonic process, fyzikální či chemický proces, při němž volná energie systému roste (ΔG > 0); děj tedy z termodynamického hlediska nemůže samovolně probíhat a musí být těsně propojen s exergonickým dějem, který mu energii dodává. K běžným endergonickým procesům patří např.:
| |
proces exergonickýangl. exergonic process, fyzikální či chemický proces, při němž volná energie systému klesá (ΔG < 0); děj tedy z termodynamického hlediska může probíhat samovolně (srov. endergonický děj). | |
proces izokratickýangl. isocratic process, laboratorní proces, při němž se nemění složení kapalného prostředí. Pojmu se užívá především při popisu chromatografických separací. Mezi izokratické procesy patří gelová chromatografie a klasická rozdělovací
chromatografie, kde se během procesu nemění složení mobilní fáze. | |
proenzymytéž zymogeny, angl. proenzymes nebo zymogens, neaktivní přímé
prekursory enzymů, které jsou do aktivní formy převáděny cíleným (specifickým) proteolytickým štěpením. Například trávicí enzymy jsou v buňkách syntetizovány jako proenzymy (trypsinogen, chymotrypsinogen, pepsinogen, prochymosin, proelastasa) a aktivovány až po vyloučení do trávicího traktu, enzymy podílející se na vzniku a odbourávání fibrinového gelu jsou v krevní plasmě přítomny ve formě prekursorů (prothrombinu, plasminogenu) a aktivují se až v případě potřeby. Bývají označovány buď předponou pro- nebo koncovkou -gen. | |
prochiralitaangl. prochirality, vlastnost některých achirálních molekul. Prochirální molekula může podlehnout enzymové přeměně tak, že dvě chemicky ekvivalentní skupiny vázané na jednom centru (prochirálním uhlíkovém atomu) mohou reagovat odlišně; enzym je může díky své stereospecifitě rozlišit. Známým příkladem je citronová kyselina, z níž se v průběhu citrátového cyklu odštěpují (jako CO2) ty uhlíkové atomy, které byly součástí molekuly oxalacetátu, z níž citrát vznikl; přitom acetátové skupiny citrátu (−CH2−COO−) jsou strukturně zcela ekvivalentní (srov. chiralita). Na obr. je patrno, jak se hydroxylová skupina působením akonitasy posunuje "dolů", ač by se stejně dobře mohla posunout "nahoru". 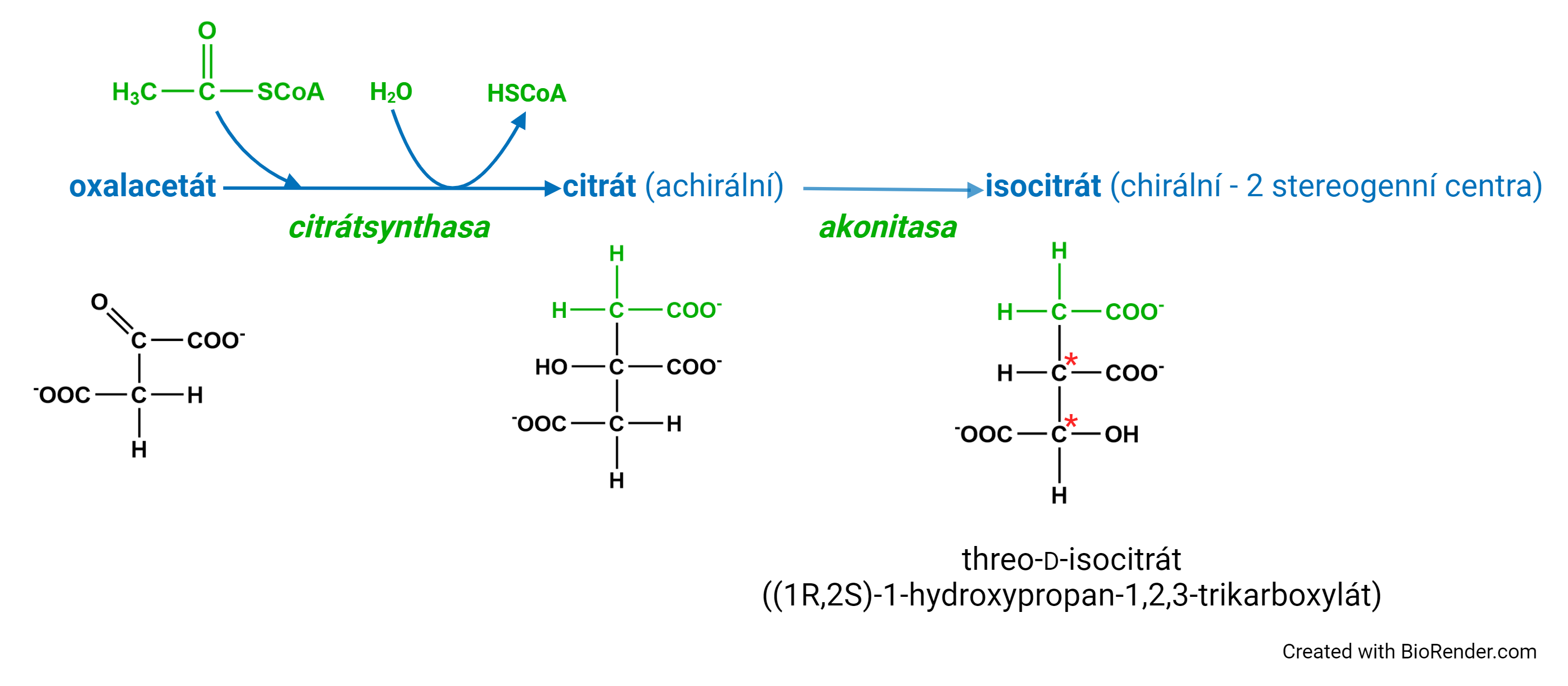 | |
prokaryotaangl. prokaryotes (řecky pro = před, karyon = jádro), jednobuněčné organismy, jejichž genetický materiál (DNA) je lokalizován v určité části buňky (v nukleoidu) a není od cytosolu oddělen membránou (viz organismy – rozdělení podle vnitřní organizace buněk). Patří sem především bakterie, archaebakterie a cyanobakterie (přesněji eubakterie). | |
prolinPro nebo P, angl. proline, hydrofobní proteinogenní neesenciální glukogenní 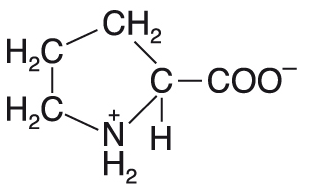 aminokyselina. Prolin je jediná proteinogenní aminokyselina, která nemá primární (volnou) aminoskupinu, ale má "postranní řetězec" v podobě cyklu; jde tedy o sekundární amin. Díky své zvláštní chemické struktuře má i specifické možnosti při vytváření prostorové struktury bílkovin. Nepodílí se na výstavbě α-helixu, ale velmi často jej nacházíme v různých ohybech peptidového řetězce. Peptidová vazba, na níž se podílí dusíkový atom prolinu, bývá v bílkovinách v konfiguraci cis. Spolu se svým posttranslačním derivátem, hydroxyprolinem, je hojně zastoupen v kolagenu. Jeho prekursorem je glutamová kyselina, jejíž cyklizací a redukcí vzniká L-prolin. Odbourává se přes glutamát na 2-oxoglutarát. aminokyselina. Prolin je jediná proteinogenní aminokyselina, která nemá primární (volnou) aminoskupinu, ale má "postranní řetězec" v podobě cyklu; jde tedy o sekundární amin. Díky své zvláštní chemické struktuře má i specifické možnosti při vytváření prostorové struktury bílkovin. Nepodílí se na výstavbě α-helixu, ale velmi často jej nacházíme v různých ohybech peptidového řetězce. Peptidová vazba, na níž se podílí dusíkový atom prolinu, bývá v bílkovinách v konfiguraci cis. Spolu se svým posttranslačním derivátem, hydroxyprolinem, je hojně zastoupen v kolagenu. Jeho prekursorem je glutamová kyselina, jejíž cyklizací a redukcí vzniká L-prolin. Odbourává se přes glutamát na 2-oxoglutarát. | |
promotorangl. promotor, charakteristická sekvence DNA, na kterou se může navázat RNA-polymerasa či jiná složka transkripčního aparátu a zahájit tak transkripci genu, jehož je promotor součástí. | |
pro-proteinyangl. proproteins, bílkoviny, které jsou syntetizovány v biologicky neaktivní formě a teprve následným proteolytickým štěpením jsou převáděny do formy aktivní (srov. proenzymy). Známými příklady pro-proteinů, nepatřících mezi enzymy, jsou fibrinogen, prokolagen a proinsulin, z nichž štěpením vznikají aktivní proteiny fibrin (monomer), kolagen a insulin. | |
pro-sekvenceangl. prosequence, úsek polypeptidového řetězce, který blokuje aktivitu daného proteinu a v procesu aktivace je odštěpován (srov. pro-proteiny). Často bývá pro-sekvence umístěna na N-konci řetězce, zdaleka to však není pravidlem (viz aktivace insulinu). | |
prostaglandinyangl. prostaglandins, oxidované deriváty arachidonové kyseliny (viz eikosanoidy), působící jako tkáňové hormony, a to zejména na hladké svaly, nervové buňky, cévní systém a reprodukční aparát. Jsou syntetizovány ve většině buněk. Některá běžná léčiva (acetylsalicylová kyselina, ibuprofen) působí jako inhibitory cyklooxygenasy (EC 1.14.99.1), klíčového enzymu syntézy prostaglandinů. Za objevy týkající se prostaglandinů a příbuzných biologicky aktivních látek získali roku 1982 S. K. Bergström, B. I. Samuelsson a J. R. Vane Nobelovu cenu. 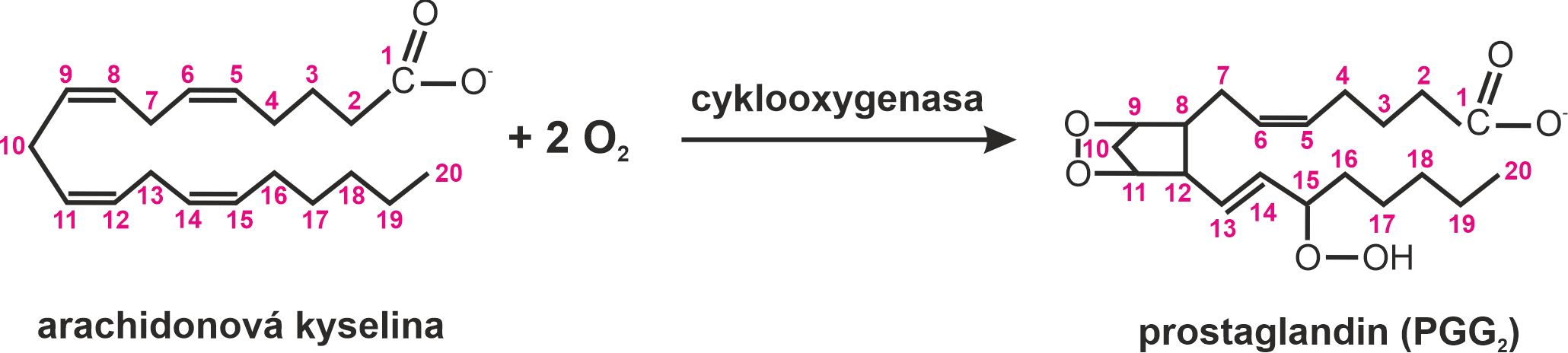 | |
proteasomtéž proteosom, angl. proteasome, bílkovinný komplex v cytosolu eukaryotních buněk, který selektivně proteolyticky degraduje (EC 3.4.25.1) bílkovinné molekuly označené
ubiquitinem. Při této exergonické reakci se štěpí ATP a získaná energie je využita pro rozbalení (isomeraci) štěpené bílkoviny (EC 5.6.1.5). | |
proteasyangl. proteases, enzymy ze třídy
hydrolas (EC 3.4.-.-), které štěpí peptidové vazby v bílkovinách. Neexistuje jasné rozhraní mezi proteasami a peptidasami. Podle katalytických skupin v aktivním místě dělíme proteasy na serinové (rozhodující je hydroxylová skupina serinu, např. trypsin, thrombin), aspartátové (skupina –COO-, např. pepsin), cysteinové (–SH, papain) a metaloproteasy (často Zn2+, karboxypeptidasa). | |
proteinasyangl. proteinases, podskupina
proteas (EC 3.4.21.-; EC 3.4.22.-; EC 3.4.23.-; EC 3.4.24.-) štěpící bílkoviny uvnitř peptidového řetězce (viz endopeptidasy). Protože většina proteas jsou endopeptidasy, používá se někdy pojem proteinasa místo obecnějšího proteasa. | |
proteinfosfátfosfatasytéž fosfoproteinfosfatasy, někdy méně přesně proteinfosfatasy, angl. phosphoprotein phosphatases, (EC 3.1.3.16), enzymy ze třídy
hydrolas, které odštěpují fosfátovou skupinu z bílkovin (viz fosfatasy). Podílejí se na jednom z nejdůležitějších způsobů regulace aktivity enzymů, spočívajícím na jejich chemické modifikaci připojením (viz proteinkinasy) a odštěpením fosfátové skupiny. Za objevy týkající se fosforylace bílkovin jakožto regulačního mechanismu získali v roce 1992 E. H. Fischer a E. G. Krebs Nobelovu cenu. | |
proteinkinasyangl. proteinkinases, enzymy ze třídy
transferas (viz kinasy), které fosforylují bílkoviny přenosem fosfátové skupiny z ATP. Modifikují postranní řetězce serinu, threoninu (Ser/Thr-proteinkinasy, EC 2.7.11.-), tyrosinu (Tyr-proteinkinasy, EC 2.7.10.-) nebo méně často histidinu (His-proteinkinasy, EC 2.7.13.-). Srov. proteinfosfátfosfatasy. | |
proteiny asociované s mikrotubulyzkr. MAPs, angl. Microtubule-Associated Proteins, bílkoviny regulující sestavování a stabilitu mikrotubulů. | |
proteiny konzervativnítéž konzervované, angl. conservative proteins, bílkoviny, jejichž pořadí aminokyselin se během evoluce ve srovnání s ostatními bílkovinami jen velmi málo mění. Vyplývá z toho, že jejich struktura je tak dokonale přizpůsobena jejich funkci a je pro život buňky (organismu) tak zásadní, že většina změn v primární struktuře není slučitelná se životem (je letální). K známým příkladům patří histony nebo ubiquitin. | |
proteoglykanyangl. proteoglycans, glykoproteiny, v nichž hmotnostně převažuje sacharidová složka. Významnými představiteli této skupiny jsou komplexy tvořené mukopolysacharidy a bílkovinami v extracelulárním prostoru pojivové tkáně. | |
proteolýzaangl. proteolysis, hydrolytické štěpení
peptidových vazeb bílkovin. Termín je používán v několika souvislostech:
| |
proteomikaangl. proteomics, obor biochemie snažící se identifikovat bílkoviny (nejlépe všechny), které jsou v dané buňce (tkáni) přítomny, a vysvětlit jejich funkci za daných fyziologických nebo patologických podmínek. Dominantními metodami proteomiky jsou dvojrozměrná elektroforéza (obvykle spojení isoelektrické fokusace a SDS-PAGE) a hmotnostní spektrometrie. Za vývoj nedestruujících ionizačních metod pro hmotnostní spektrometrii biologických makromolekul získali roku 2002 J. B. Fenn a K. Tanaka Nobelovu cenu. | |
proteosyntézatéž exprese genu, angl. proteosynthesis, biosyntéza bílkovin, komplexní proces sestávající z několika kroků:
| |
prothrombin | |
protilátkytéž imunoglobuliny, angl. antibodies, proteiny v krevní plasmě schopné vázat
antigeny. Protilátky, které se vážou na jeden (makromolekulární) antigen, mohou být polyklonální (vážou se na více antigenních determinant antigenu) nebo monoklonální (vážou se na jedinou determinantu). Monoklonální protilátky se získávají hybridomovými technologiemi. Polyklonální protilátka je vlastně směsí více monoklonálních protilátek zaměřených proti jednomu antigenu. Za popis struktury protilátek získali G. M. Edelman a R. R. Porter v roce 1972 Nobelovu cenu; v roce 1984 ji za objevy umožňující přípravu monoklonálních protilátek obdrželi G. J. F. Köhler a C. Milstein . | |
protoplastangl. protoplast, buňka rostlinného nebo mikrobiálního původu, zbavená
buněčné stěny. Takto upravené buňky jsou vhodným objektem pro různé modifikace genomu a fyziologické studie. | |
protoporfyriny | |
provitaminyangl. provitamins, neaktivní
prekursory vitaminů; na svou aktivní formu jsou přeměňovány chemickými modifikacemi v organismu. Nejznámějšími provitaminy jsou karotenoidy (provitaminy A) a ergosterol (provitamin D). | |
Prusiner Stanley Ben
| |

pseudogen | |
pseudoisoenzymyangl. pseudoisoenzymes, enzymy, které se vyskytují v jednom organismu, katalyzují stejnou reakci a vznikají transkripicí stejného genu; mají však jinou chemickou strukturu, protože podléhají odlišným posttranslačním (nebo kotranslačním) modifikacím (srov. isoenzymy). | |
pufrytlumivé roztoky, ve starší literatuře též nárazníky nebo ústojné roztoky, angl. buffers, vodné roztoky, jejichž pH se (ve srovnání s vodou) po přidání silné kyseliny nebo zásady mění jen málo. Mají tedy schopnost tlumit změny pH. Mírou této schopnosti je pufrační kapacita; je definována jako množství jednosytné silné kyseliny, kterou je třeba přidat k roztoku, aby pH kleslo o jednotku. V biochemii se s pufry setkáváme ve dvou souvislostech:
| |
pumpy iontovéangl. ion pumps, integrální transmembránové bílkoviny obvykle oligomerní povahy, které zajišťují selektivní aktivní transport iontů; v užším slova smyslu se tím často míní pouze primární aktivní transport, který zajišťují enzymy třídy translokas využívající ATP (EC 7.2.2.-). Nejvýznamnější iontovou pumpou je Na,K-ATPasa; podobně fungují i iontové pumpy zajišťující např. transport vodíkových protonů do prostoru žaludku (EC 7.2.2.19), čímž zde výrazně snižují hodnotu pH, nebo vápenatých iontů z cytosolu do sarkoplasmatického retikula (EC 7.2.2.10). | |
purin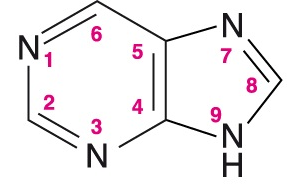 angl. purine, heterocyklická dusíkatá sloučenina, strukturní základ purinových bazí (adenin, guanin, hypoxanthin).
Za syntézu purinových derivátů obdržel roku 1902 H. E. Fischer Nobelovu cenu. angl. purine, heterocyklická dusíkatá sloučenina, strukturní základ purinových bazí (adenin, guanin, hypoxanthin).
Za syntézu purinových derivátů obdržel roku 1902 H. E. Fischer Nobelovu cenu. | |
pyranosyangl. pyranoses, cyklické formy monosacharidů, formálně | |
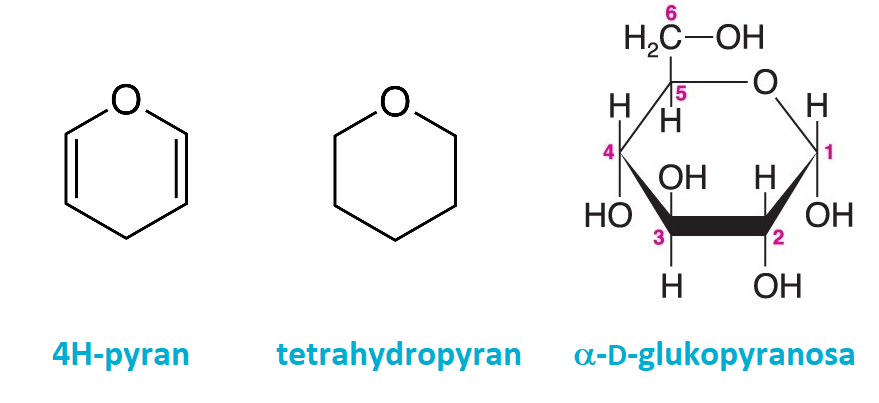 odvozené od šestičlenného heterocyklu pyranu.
odvozené od šestičlenného heterocyklu pyranu.pyridoxinangl. pyridoxine, viz vitamin B6. | |
pyrimidinangl. pyrimidine, dusíkatý heterocyklus, strukturní základ pyrimidinových bází nukleových kyselin cytosinu, thyminu a uracilu. 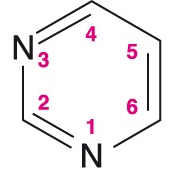 | |
pyrolysintéž pyrrolysin, Pyl nebo U, angl. pyrrolysine, vzácná bazická aminokyselina | |
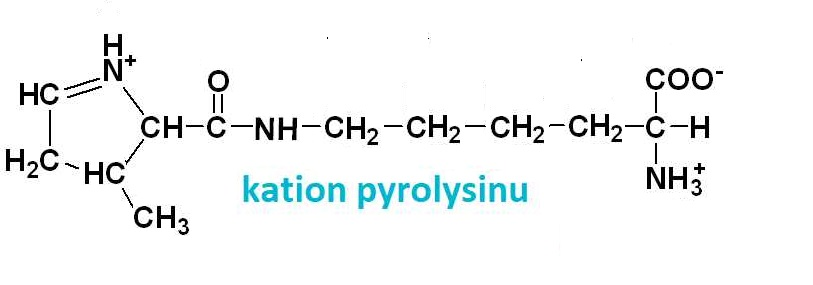 objevená r. 2002 v určitých
objevená r. 2002 v určitých pyruvátanion pyrohroznové kyseliny, angl. pyruvate, jeden z nejvýznamnějších metabolitů. 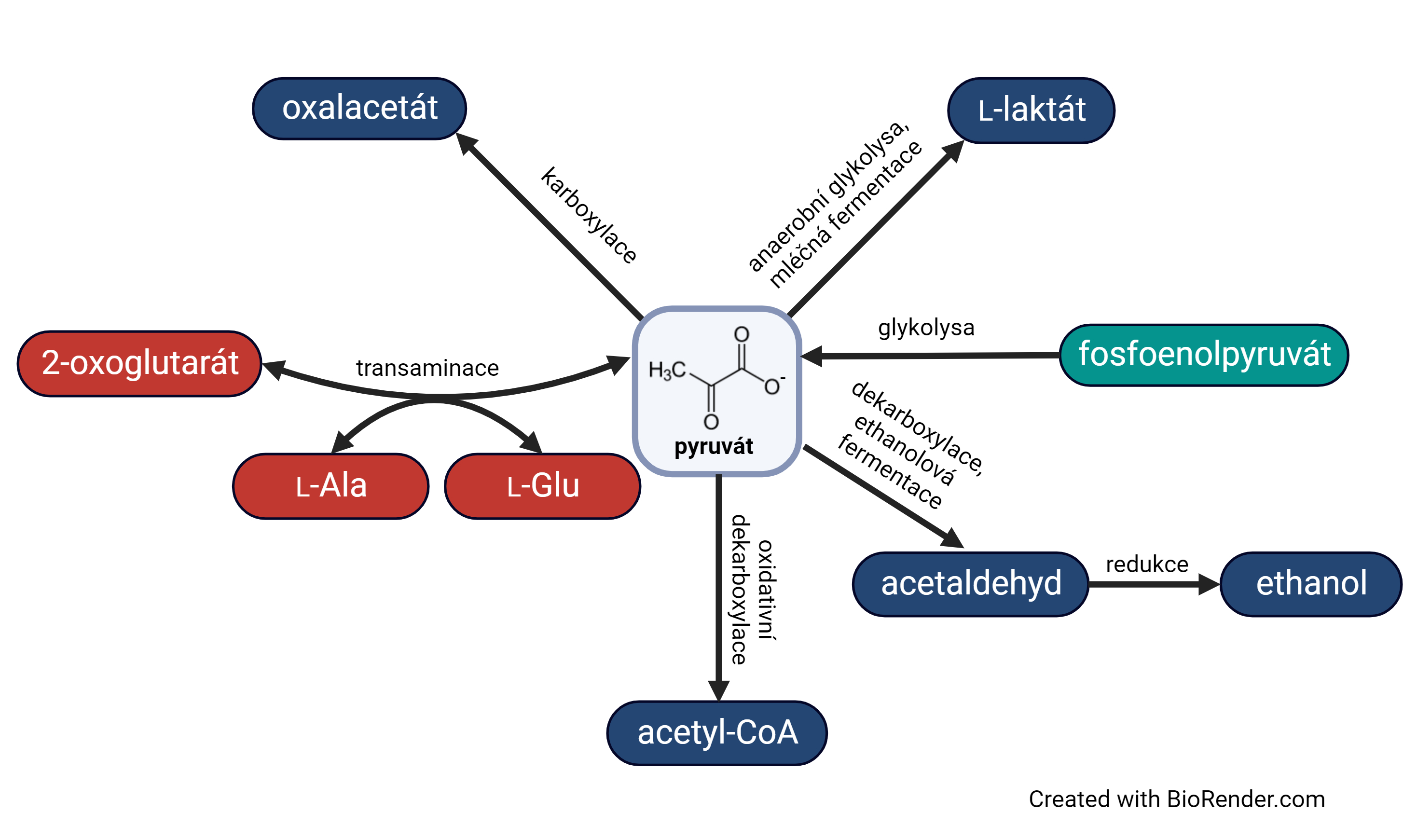 Vzniká v
cytosolu v závěrečné fázi glykolysy. Po přechodu do mitochondrie (symport spolu s H+) může být oxidačně dekarboxylován za vzniku acetyl-CoA, transaminací se může přeměňovat na aminokyselinu alanin, karboxylací přechází na oxalacetát (meziprodukt citrátového cyklu, prekursor biosyntézy glukosy), za anaerobních podmínek může být redukován na laktát nebo dekarboxylován na acetaldehyd (výchozí krok alkoholového kvašení). Vzniká v
cytosolu v závěrečné fázi glykolysy. Po přechodu do mitochondrie (symport spolu s H+) může být oxidačně dekarboxylován za vzniku acetyl-CoA, transaminací se může přeměňovat na aminokyselinu alanin, karboxylací přechází na oxalacetát (meziprodukt citrátového cyklu, prekursor biosyntézy glukosy), za anaerobních podmínek může být redukován na laktát nebo dekarboxylován na acetaldehyd (výchozí krok alkoholového kvašení). | |