Biochemický slovník
Biochemický slovník

Procházet slovníkem pomocí tohoto rejstříku
Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
2 |
|---|
2-oxotéž glutamát, Glu nebo E, angl. glutamic acid nebo glutamate, proteinogenní 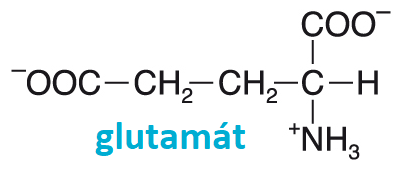 glukogenní neesenciální aminokyselina kyselé povahy. Volná se uplatňuje v mnohých reakcích, zejména při přenosu dusíkového atomu (transaminace). Velmi důležitá je dehydrogenace glutamátu doprovázená odštěpením amoniaku (viz glutamátdehydrogenasa). Je prekursorem biosyntézy neesenciálních aminokyselin glutaminu a prolinu. Vyrábí se průmyslově ve velkém množství, její monosodná sůl (glutamát sodný) se přidává do potravin. Glutamová kyselina je nejdůležitějším excitačním (budivým) neurotransmiterem v centrální nervové soustavě (až 75 % excitace) a v sítnici. Její dekarboxylací vzniká tlumivý neurotransmiter GABA (viz kyselina γ-aminomáselná). glukogenní neesenciální aminokyselina kyselé povahy. Volná se uplatňuje v mnohých reakcích, zejména při přenosu dusíkového atomu (transaminace). Velmi důležitá je dehydrogenace glutamátu doprovázená odštěpením amoniaku (viz glutamátdehydrogenasa). Je prekursorem biosyntézy neesenciálních aminokyselin glutaminu a prolinu. Vyrábí se průmyslově ve velkém množství, její monosodná sůl (glutamát sodný) se přidává do potravin. Glutamová kyselina je nejdůležitějším excitačním (budivým) neurotransmiterem v centrální nervové soustavě (až 75 % excitace) a v sítnici. Její dekarboxylací vzniká tlumivý neurotransmiter GABA (viz kyselina γ-aminomáselná).  | |
2-oxoglutarátangl. 2-oxoglutarate, ve starší terminologii α-ketoglutarát, anion 2-oxoglutarové kyseliny, akceptor aminoskupin při transaminacích (viz kyselina glutamová), meziprodukt citrátového cyklu, kde podléhá 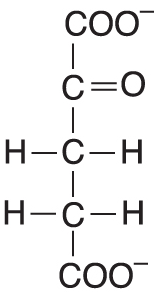 oxidační dekarboxylaci (viz multienzymové jednotky). oxidační dekarboxylaci (viz multienzymové jednotky). | |
5 |
|---|
5'-fosforibosyl-1'-difosfátangl. 5’-phosphoribosyl-1’-pyrrophosphate, zkr. PRPP, významný meziprodukt biosyntézy purinových bází, ale také (u rostlin a mikroorganismů) biosyntézy 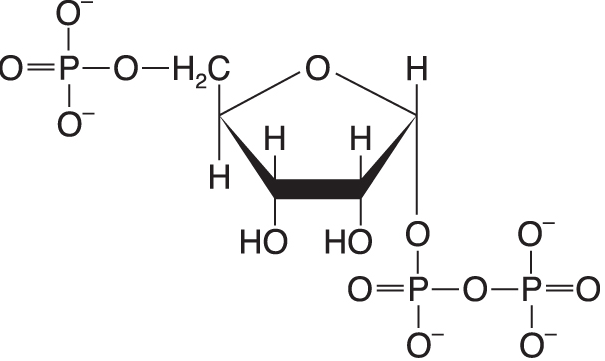 pro člověka esenciálních aminokyselin histidinu a tryptofanu. pro člověka esenciálních aminokyselin histidinu a tryptofanu. | |
Α |
|---|
α-fetoproteinangl. α-fetoprotein, zkr. AFP, glykoprotein tvořený játry lidského plodu a vyskytující se v malém množství v plodové vodě, z níž přestupuje do mateřské krve. V dospělosti je nahrazen sérovým albuminem. Přítomnost α-fetoproteinu v séru dospělých je indikátorem přítomnosti některých nádorů, především karcinomu jater. Hladina α-fetoproteinu v séru těhotných žen a v plodové vodě se klinicky stanovuje ve snaze o včasnou diagnostiku některých vrozených vad plodu. | |
α-helixangl. α-helix, časté periodické prostorové uspořádání polypeptidových řetězců v bílkovinách. Jeden závit pravotočivé šroubovice tvoří 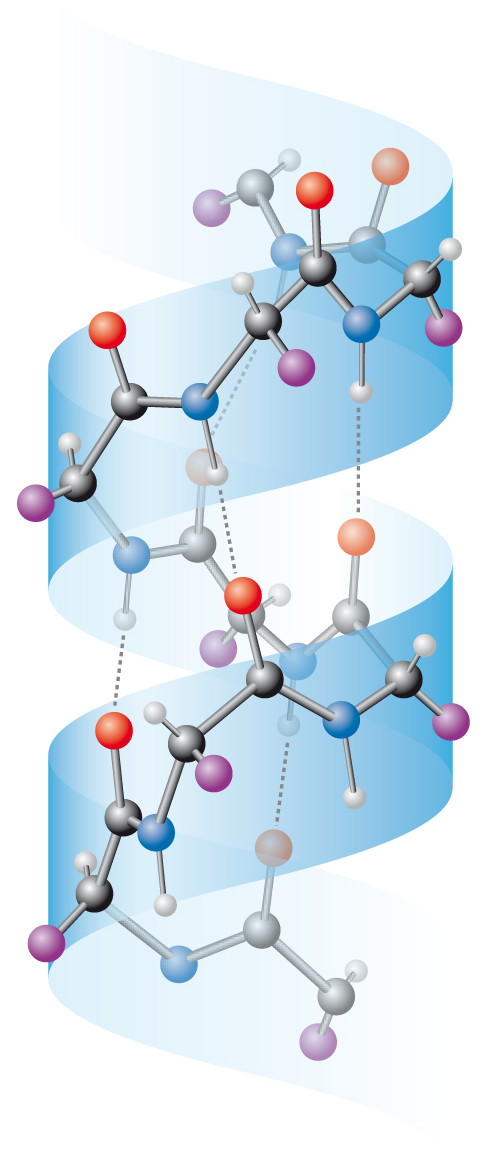 3,6 aminokyselinových zbytků. Tento typ helixu je stabilizován vnitrořetězcovými vodíkovými vazbami spojujícími skupiny CO a NH v sousedních závitech šroubovice. U některých globulárních bílkovin (např. hemoglobinu) může až 75 % peptidového řetězce zaujímat tuto strukturu. V bílkovinách, zejména pak strukturních (např. kolagenech) mohou existovat i jiné typy helixů; pro ně se samozřejmě neužívá označení α. 3,6 aminokyselinových zbytků. Tento typ helixu je stabilizován vnitrořetězcovými vodíkovými vazbami spojujícími skupiny CO a NH v sousedních závitech šroubovice. U některých globulárních bílkovin (např. hemoglobinu) může až 75 % peptidového řetězce zaujímat tuto strukturu. V bílkovinách, zejména pak strukturních (např. kolagenech) mohou existovat i jiné typy helixů; pro ně se samozřejmě neužívá označení α. | |
Β |
|---|
β-oxidace mastných kyselintéž Linenova spirála, angl. fatty acids β-oxidation, katabolický proces, při němž se z aktivované nasycené
mastné kyseliny (acyl-CoA) odštěpuje dvouuhlíkatý štěp (acetyl-CoA) za vzniku mastné kyseliny o dva uhlíky kratší:
kde ETF je zkratka pro electron-transfer flavoprotein, který prostřednictvím své prostetické skupiny FAD působí jako součást postranního vstupu do dýchacího řetězce. V eukaryotních buňkách probíhá tento proces v mitochondrii. Při úplném aerobním odbourání nasycené mastné kyseliny se sudým počtem uhlíků lze počet získaných jednotek ATP vypočítat ze vztahu ATP = 12n/2 + 5(n – 2)/2 – 2, neboť úplným odbouráním acetylu v citrátovém cyklu a dýchacím řetězci se získá 12 ATP, reoxidací NADH 3 ATP, reoxidací FADH2 2 ATP a 2 jednotky ATP byly investoványdo aktivace mastné kyseliny (R–COOH + CoA + ATP → R–CO–CoA + AMP + PPi). Obdobným způsobem, byť s určitými obměnami, jsou odbourávány i mastné kyseliny nenasycené, s lichým počtem uhlíků a s rozvětvenými řetězci. Za objevy týkající se mechanismu a regulace metabolismu mastných kyselin získali roku 1964 F. Lynen a K. E. Bloch Nobelovu cenu. | |
Γ |
|---|
γ-globulinyangl. γ-globulins, frakce globulinů krevního séra. Patří mezi ně i imunoglobuliny; γ-globuliny se injekčně aplikují pro nespecifické zvýšení imunity. | |
γ-glutamyltransferasatéž gama-glutamyltransferasa (GGT, gamma-GT, EC 2.3.2.2), angl. gamma-glutamyl tranferase, enzym přenášející γ-glutamylový zbytek (např. z glutathionu) na volnou aminoskupinu aminokyseliny nebo peptidu: (5-L-glutamyl)-peptid + aminokyselina ⇌ peptide + 5-L-glutamyl-aminokyselina. Hraje významnou roli při syntéze a odbourávání glutathionu, podílí se také na detoxifikaci xenobiotik. Jedná se o membránový enzym, podílející se na transportu aminokyselin. Nachází se především v jaterních buňkách; při jaterních onemocněních bývá koncentrace její katalytické aktivity v krevním séru významně zvýšena. | |