Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
R |
|---|
ribozymyangl. ribonucleic enzymes nebo ribozymes, ribonukleové kyseliny s katalytickou aktivitou. Byly popsány především ribozymy působící na RNA (viz sestřih RNA); jistou enzymovou funkci vykazují i rRNA. Existence RNA s enzymovou aktivitou vede k domněnce, že prvotními biopolymery v evoluci mohly být nukleové kyseliny a že tedy proteiny vznikly později. Za objev katalytických vlastností RNA získali roku 1989 S. Altman a T. R. Cech Nobelovu cenu. | |
RNAribonukleová kyselina, angl. ribonucleic acid, lineární polymer ribonukleotidů (A,G,C,U), spojených fosfodiesterovými vazbami. Na rozdíl od DNA je pouze jednořetězcová. Je syntetizována procesem transkripce podle DNA, v níž je uložena informace o pořadí ribonukleotidů ve vznikající RNA (princip komplementarity bází). RNA většinou podléhá dalším úpravám (viz posttranskripční modifikace). Rozlišujeme několik základních typů RNA, které se liší prostorovou strukturou a zejména funkcí:
| |
RNA-interference
zkr. RNAi, angl. RNA interference, mechanismus, jímž jsou v buňkách likvidovány dvouřetězcové molekuly RNA (double-stranded RNA, dsRNA). Předpokládá se, že tento mechanismus vznikl jako obrana buněk proti infekci virovou dsRNA. Při využití v genovém inženýrství může být specifická dsRNA do buňky vpravena mikroinjekcí, elektroporací apod. V současnosti se tato metoda hojně využívá k cílenému útlumu exprese určitých genů (gene knock-out, gene silencing nebo RNA silencing). Využívá se toho, že vložený řetězec RNA, který je komplementární k buněčné mRNA, s ní může vytvořit dsRNA. Tato dvouřetězcová RNA je pak mechanismem RNAi likvidována a nemůže vstoupit do procesu translace.
Nejznámější mechanismus RNAi je následující: 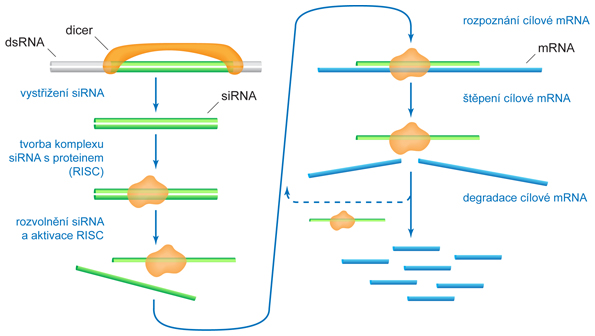 1. dsRNA je v buňce rozštěpena specifickou endonukleasou zvanou DICER. Vznikají tak krátké dvouřetězcové oligonukleotidy obsahující 20 až 25 párů bází, zvané siRNA (short-interfering RNA). 2. siRNA se naváže na specifický proteinový komplex RISC (RNA-induced silencing complex), který řetězce oddělí od sebe a na jeden z nich (antisense RNA) naváže molekulu komplementární mRNA. 3. Takto vzniklý komplex vykazuje endonukleasovou aktivitu a je schopen navázanou mRNA rozštěpit a znemožnit, aby byla použita jako templát pro translaci. Fragmenty mRNA se odpojí a celý proces (od bodu 2) se může opakovat. Tímto způsobem může jedna molekula siRNA pomoci vyhledat a inaktivovat mnoho molekul určité mRNA. 4. Specifická RNA-dependentní-RNA-polymerasa, vyskytující se u všech eukaryot kromě hmyzu a savců, může jednotlivé řetězce siRNA navázané na mRNA využít jako primery a prodloužit dvouřetězcové úseky RNA. Ty se pak stanou substrátem pro DICER (viz bod 1) a cyklus se opakuje. Jedna molekula antisense RNA (nebo dsRNA) tak může způsobit likvidaci velkého počtu molekul mRNA.
Dalším typem antisense RNA vyskytujícím se přirozeně v buňkách je pre-micro RNA. Tyto molekuly jsou syntetizovány podle určitých nekódujících úseků DNA a podílejí se na regulaci exprese určitých genů. Obsahují vlásenkovité struktury, které připomínají dsRNA, a jsou proto štěpeny DICERem za vzniku mikro RNA (miRNA), která se váže na RISC. Mohou zabránit translaci mRNA, která je k jednomu z řetězců komplementární. Za objev RNA-interference (tlumení exprese genů dvouřetězcovou RNA) získali roku 2006 A. Z. Fire a C. C. Mello Nobelovu cenu. | |
RNA-polymerasapřesněji DNA-dependentní-RNA-polymerasa, EC 2.7.7.6, angl. RNA polymerase, enzym ze třídy
transferas katalyzující biosyntézu RNA v procesu transkripce: (NMP)n + NTP → (NMP)n+1 + PPi , kde (NMP)n je prodlužující se oligoribonukeotid a NTP ribonukleosidtrifosfát. RNA-polymerasa je tvořena několika podjednotkami s některými specifickými funkcemi; např. σ-podjednotka u prokaryot zajišťuje vazbu enzymu na promotor, a tím iniciaci transkripce. | |
Robinson Robert | |
Rodbell Martin
| |

Rose Irwinamerický biochemik ( | |

rostliny C3, C4 a CAMangl. C3, C4 a CAM plants, rozlišení rostlin podle způsobu fixace
CO2 při fotosyntéze. U rostlin C3 reaguje CO2 s ribulosa-1,5-bisfosfátem za katalýzy enzymem RUBISCO a prvním detekovatelným produktem je tříuhlíkatý (proto C3) meziprodukt glukogenese 3-fosfoglycerát. Do této skupiny patří většina rostlin mírného pásma. Nevýhodou tohoto způsobu fixace CO2 je to, že v důsledku nízkého parciálního tlaku CO2 v atmosféře a vysokého parciálního tlaku O2 zde probíhá poměrně intenzivní fotorespirace. U rostlin C4 je primárním akceptorem CO2 fosfoenolpyruvát a vzniká čtyřuhlíkatý oxalacetát (viz cyklus C4-rostlin); mezi C4-rostliny patří mnohé tropické rostliny, z našich pak zejména kukuřice. Rostliny CAM (Crassulacean Acid Metabolism – metabolismus kyselin u tučnolistých) v noci fixují CO2 podobným způsobem jako C4-rostliny, uchovávají malát ve vakuolách a ve dne, kdy je dostatek světla a tedy i ATP a NADPH, realizují Calvinův cyklus; je tedy u nich časově oddělena fixace CO2 od jeho využití. Tyto rostliny, např. kaktusovité nebo agávovité, jsou adaptovány na horké a suché podnebí tím, že během dne mohou mít uzavřené průduchy a brání se tak vysychání. | |
Rothman James Edward
| |
 americký buněčný biolog (nar. 1950), Nobelova cena (za fyziologii a lékařství, 2013) za studii regulace
americký buněčný biolog (nar. 1950), Nobelova cena (za fyziologii a lékařství, 2013) za studii regulace rovnice Goldmanova
též Goldmanova–Hodgkinova–Katzova rovnice, angl. Goldman equation, vztah umožňující vypočítat membránový potenciál (Δφ) pomocí intracelulárních (i) a extracelulárních (e) koncentrací nejdůležitějších iontů (Na+, K+, Cl−) a pomocí koeficientůbpropustností P těchto iontů membránou:
\( \Delta \phi= \frac{RT}{F} ln \frac{ P_{Na} [ Na^{+}]_e +P_{K} [ K^{+}]_e + P_{Cl} [ Cl^{-} ]_i }{P_{Na} [ Na^{+}]_i +P_{K} [ K^{+}]_i + P_{Cl} [ Cl^{-} ]_e} \) Vzhledem k tomu, že koncentrace iontů na obou stranách membrány se příliš nemění, jsou prudké změny membránového potenciálu, charakteristické např. pro vedení nervového vzruchu, způsobeny náhlými změnami pasivní propustnosti iontů (viz pasivní transport, iontové kanály). | |