Biochemický slovník
Biochemický slovník

Procházet slovníkem pomocí tohoto rejstříku
Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
F |
|---|
fotofosforylaceangl. photophosphoralytion, syntéza
ATP při fotosyntéze, probíhající podle rovnice ADP + Pi → ATP + H2O. Jedná se o typ membránové fosforylace, kdy proton-motivní síla je generována na úkor zachycené světelné energie. Rozlišují se dva způsoby provedení tohoto děje:
| |
fotolýza vodytéž Hillova reakce, angl. water photolysis, rozklad vody při
fotosyntéze u sinic, řas a vyšších rostlin. Po absorpci světelného kvanta se z molekuly chlorofylu v reakčním centru fotosystému II uvolní elektron, který je použit v dalších reakcích fotosyntézy; tento elektron je posléze nahrazen elektronem z molekuly vody a molekulový kyslík se uvolňuje. Obvykle se tato reakce vyjadřuje v symbolické podobě: 2 H2O → O2 + 4 H+ + 4 e-. Vzhledem k tomu, že uvolněný elektron ve skutečnosti nahrazuje elektron, použitý pro redukci NADP, je lépe celkovou bilanci děje vyjádřit jako: H2O + NADP+→ ½ O2 + NADPH + H+ (viz fotoredukce). Atom kyslíku ve vodě zde působí, v rozporu se vší chemickou zkušeností (kyslík se přece považuje za prototyp oxidačního činidla), jako redukční činidlo. Fotolýza vody je jediným zdrojem molekulového kyslíku na Zemi. | |
fotoredukceangl. photoreduction, redukce
NADP+ při fotosyntéze: NADP+ + 2 e- + H+ → NADPH. V celkové bilanci fotosyntézy může být zdrojem elektronů voda (viz fotolýza vody) nebo jiný zdroj elektronů (viz oxygenní a anoxygenní fotosyntéza). | |
fotorespiraceangl. photorespiration, světelné dýchání rostlin. Enzym ribulosa-1,5-bisfosfátkarboxylasa/oxygenasa (viz RUBISCO) má schopnost 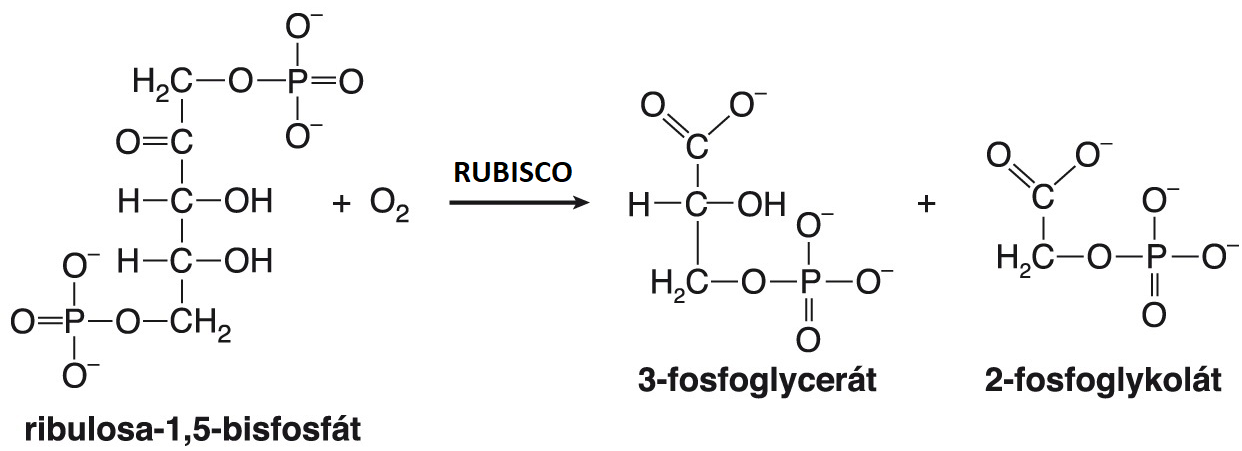 vázat na substrát (ribulosa-1,5-bisfosfát) oxid uhličitý, ale také kyslík. K fotorespiraci dochází oxygenasovou aktivitou tohoto enzymu. Po navázání kyslíku vzniká nestálý meziprodukt s pěti atomy uhlíku, který se rozpadá na dva produkty, jeden se třemi a druhý se dvěma atomy uhlíku. Tříuhlíkatý produkt (3-fosfoglycerát) vstupuje do reakcí Calvinova cyklu, dvouuhlíkatý (2-fosfoglykolát) je dále metabolizován v peroxisomu na glyoxylát, který může přijmout aminoskupinu a stát se tak prekursorem syntézy glycinu a následně serinu. Většina glyoxylátu je však katabolizována až na CO2, a proto fotorespirace působí značný pokles produkce sacharidů. Dochází k ní především u C3-rostlin (viz rostliny C3, C4 a CAM), kde může způsobit ztrátu 20 až 40 % primárně zachyceného CO2. | |
fotosyntézaangl. photosynthesis, proces zahrnující zachycení energie světelného záření, její uložení ve formě energie chemických vazeb a její následné využití pro přeměnu anorganických látek (zejména
CO2) na látky organické. Fotosyntéza je nejdůležitější biochemický proces na Zemi. Probíhá v zelených částech rostlin, fototrofních bakteriích a řasách. U eukaryot je lokalizována v chloroplastech, specializovaných organelách obsahujících chlorofyl. Ve vnitřních membránách chloroplastů (thylakoidech) jsou zakotveny dva typy fotosystémů absorbujících světelnou energii. Takto získaná energie je ve světlé fázi fotosyntézy přímo využita k redukci NADP+ na NADPH (viz fotoredukce), ale také uložena do formy proton-motivní síly a následně do chemické struktury ATP (viz fotofosforylace). Tyto látky jsou nezbytné pro následující temnou fázi fotosyntézy, kde se zabudovává CO2 (v Calvinově cyklu), ale také anorganický dusík a síra do organických sloučenin. Podle prvních kroků fixace CO2 dělíme rostliny na C3, C4 a CAM (viz rostliny C3, C4 a CAM). | |
fotosyntéza oxygenní a anoxygenníangl. oxygenic and anoxygenic fotosynthesis, dvě varianty světlé fáze fotosyntézy. Při oxygenní fotosyntéze je donorem elektronů pro fotoredukci voda (resp. její kyslík, viz fotolýza vody), při anoxygenní nějaký jiný donor elektronů. Anoxygenní fotosyntéza je vývojově starší forma, v současnosti přetrvávající pouze u některých bakterií, které mohou být fotolithotrofní nebo fotoorganotrofní (viz organismy – rozdělení podle způsobu výživy). Donorem vodíku pro fotoredukci je u fotolithotrofních bakterií nejčastěji sulfan (produkují síru), u fotoorganotrofních bakterií organická látka, např. 2-propanol (produkují aceton) nebo mastná kyselina. Konečným akceptorem vodíku je vždy oxid uhličitý, který je, jako při každém fotosyntetickém procesu, redukován a zabudován do organických molekul, především do sacharidů. Protože při anoxygenní fotosyntéze mají donory elektronů nižší redoxní potenciál (jsou slabšími oxidačními činidly) než kyslík a uvolňují tedy elektron snáze, mohou fotosyntetizující bakterie vystačit s jediným fotosystémem. | |
fotosystémyangl. photosystems, membránové strukturní jednotky zachycující a využívající světelnou energii, též
kvantosomy. Jsou uloženy ve velkém množství ve vnitřních membránách chloroplastů (v thylakoidech). Rozlišujeme dva typy, fotosystém I a fotosystém II (PS I a PS II). Jsou tvořeny anténním komplexem (LHC) a reakčním centrem (RC). Molekuly LHC absorbují světelnou energii a přenášejí ji do RC, kde je přítomen chlorofyl a, vázaný na specifický protein. Při excitaci se dramaticky sníží redoxní potenciál tohoto chlorofylu, který se tak stává silným redukčním činidlem a je schopen přenést svůj elektron na molekulu příslušného přenašeče. Absorpční maximum chlorofylu a v PS I je 700 nm, v PS II 680 nm, proto jsou někdy označovány jako fotosystém P700 a P680. | |
fragmenty Okazakihoangl. Okazaki fragments, polydeoxyribonukleotidy, které jsou syntetizovány při
replikaci jakožto úseky zpožďujícího se řetězce, aby byly posléze spojeny DNA-ligasou a vytvořily jediné vlákno. Každý úsek se syntetizuje jako samostatná jednotka včetně iniciace pomocí RNA-polymerasy. U prokaryot mají Okazakiho fragmenty délku 1 000 až 2 000 deoxyribonukleotidů, u eukaryot jsou mnohem kratší (100 až 200 deoxyribonukleotidů). | |
fruktosatéž ovocný cukr, angl. fructose, nejrozšířenější ketohexosa, nejsladší 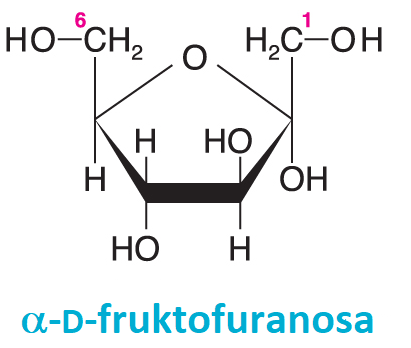 přírodní cukr. Jako všechny významné monosacharidy existuje v přírodě téměř výhradně v konfiguraci D; otáčí však rovinu lineárně polarizovaného světla vlevo, proto se pro ni používá tradiční synonymum levulosa. Fruktosa se vyskytuje v různých rostlinných šťávách a v medu. Je rychleji stravitelná než glukosa. Disacharid tvořený fruktosou a glukosou je sacharosa. Fruktosa (resp. její aktivované formy, fruktosa-6-fosfát a fruktosa-1,6-bisfosfát) jsou meziprodukty glykolysy, glukogenese, pentosového a Calvinova cyklu. Fruktosa-2,6-bisfosfát je důležitým regulačním metabolitem. přírodní cukr. Jako všechny významné monosacharidy existuje v přírodě téměř výhradně v konfiguraci D; otáčí však rovinu lineárně polarizovaného světla vlevo, proto se pro ni používá tradiční synonymum levulosa. Fruktosa se vyskytuje v různých rostlinných šťávách a v medu. Je rychleji stravitelná než glukosa. Disacharid tvořený fruktosou a glukosou je sacharosa. Fruktosa (resp. její aktivované formy, fruktosa-6-fosfát a fruktosa-1,6-bisfosfát) jsou meziprodukty glykolysy, glukogenese, pentosového a Calvinova cyklu. Fruktosa-2,6-bisfosfát je důležitým regulačním metabolitem. | |
furanosyangl. furanoses, cyklické formy monosacharidů, formálně | |
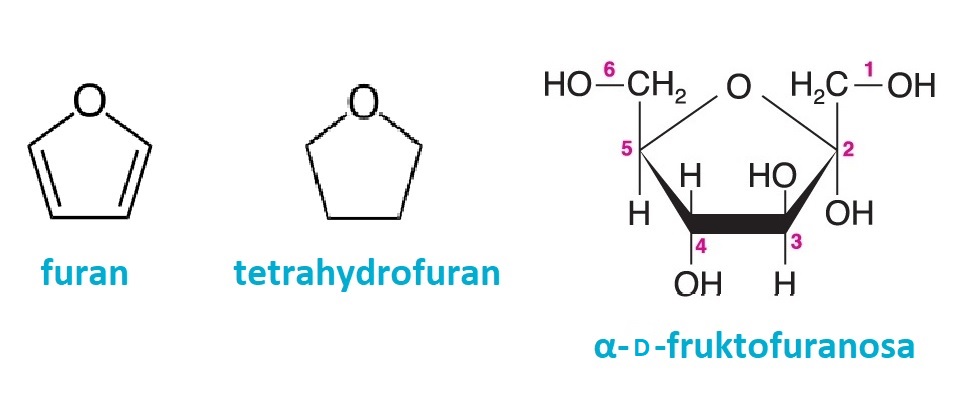 odvozené od pětičlenného heterocyklu furanu.
odvozené od pětičlenného heterocyklu furanu.