Biochemický slovník
Biochemický slovník

Procházet slovníkem pomocí tohoto rejstříku
Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
F |
|---|
fosfoenolpyruvátpřesněji 2-fosfoenolpyruvát, zkr. PEP, angl. phosphoenolpyruvate, důležitá makroergická sloučenina, 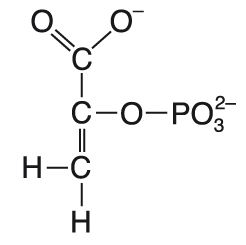 meziprodukt glykolysy a glukogenese, akceptor CO2 v cyklu C4-rostlin. Svůj fosfátový zbytek může předat ADP za vzniku ATP a pyruvátu (substrátová fosforylace). meziprodukt glykolysy a glukogenese, akceptor CO2 v cyklu C4-rostlin. Svůj fosfátový zbytek může předat ADP za vzniku ATP a pyruvátu (substrátová fosforylace). | |
fosfofruktokinasaangl. phosphofructokinase, EC 2.7.1.11, enzym zodpovědný za regulaci glykolysy u savců. Katalyzuje fosforylaci fruktosa-6-fosfátu: fruktosa-6-fosfát + ATP → fruktosa-1,6- bisfosfát + ADP. Jaterní enzym je allostericky inhibován meziprodukty glykolysy a citrátového cyklu (fosfoenolpyruvátem a citrátem) a především ATP (indikátor "energetického dostatku"); jejich vazbou je enzym stabilizován v neaktivním stavu a tím je snížena jeho afinita k fruktosa-6-fosfátu. ADP a AMP (indikátory "energetické nouze") naopak stabilizují enzym v aktivním stavu a zvyšují tak afinitu k substrátům (fruktosa-6-fosfát a ATP). Tento enzym je také regulován hormonálně prostřednictvím insulinu. | |
fosfolipasyangl. phospholipases, skupina hydrolas, které štěpí fosfolipidy, zejména fosfatidy. Podle místa štěpení fosfatidů je  dělíme na fosfolipasu A1 (EC 3.1.1.32) odštěpující mastné kyseliny v poloze 1 glycerolu (např. palmitovou kyselinu z fosfolipidu na obr.), fosfolipasu A2 (EC 3.1.1.4, odštěpující olejovou kyselinu v poloze 2 ), fosfolipasu C (EC 3.1.4.3) štěpící vazbu mezi fosfátem a diacylglycerolem (na obr., vzniká IP3), a fosfolipasu D (EC 3.1.4.4), odštěpující alkohol z fosfatidové kyseliny (na obr., vzniká inositol-4,5-bisfosfát). Běžně se užívají zkratky PLA1, PLA2, PLC a PLD. Fosfolipasy se podílejí na odbourávání fosfolipidů, některé z nich, zejména PLC a PLD, hrají významnou roli při přenosu signálu (viz fosfatidylinositoly). dělíme na fosfolipasu A1 (EC 3.1.1.32) odštěpující mastné kyseliny v poloze 1 glycerolu (např. palmitovou kyselinu z fosfolipidu na obr.), fosfolipasu A2 (EC 3.1.1.4, odštěpující olejovou kyselinu v poloze 2 ), fosfolipasu C (EC 3.1.4.3) štěpící vazbu mezi fosfátem a diacylglycerolem (na obr., vzniká IP3), a fosfolipasu D (EC 3.1.4.4), odštěpující alkohol z fosfatidové kyseliny (na obr., vzniká inositol-4,5-bisfosfát). Běžně se užívají zkratky PLA1, PLA2, PLC a PLD. Fosfolipasy se podílejí na odbourávání fosfolipidů, některé z nich, zejména PLC a PLD, hrají významnou roli při přenosu signálu (viz fosfatidylinositoly). | |
fosfolipidyangl. phospholipids, lipidy obsahující fosfátový zbytek; biologicky velmi významná skupina složených
lipidů, tvořících základní složku biologických membrán. Chemicky to jsou zejména:
| |
fosfoproteinyangl. phosphoproteins, složené bílkoviny, jejichž prostetickou skupinou je kyselina fosforečná (resp. anorganický fosfát), většinou vázaná na hydroxylové skupiny postranních řetězců serinu, threoninu nebo tyrosinu esterovou vazbou. Dočasná fosforylace enzymů patří k nejdůležitějším způsobům regulace jejich aktivity. Trvale fosforylované bílkoviny jsou obsaženy ve velkém množství v mléce (kasein) a ve vejcích (ovalbumin, fosfovitin); mezi trvalé fosfoproteiny patří i pepsin. | |
fosforylaceangl. phosphorylation, připojení zbytku kyseliny fosforečné, lépe řečeno enzymově katalyzovaná tvorba jejích esterů či anhydridů. Nejčastějším způsobem fosforylace je přenos zbytku kyseliny fosforečné z
ATP na substrát (např. ATP + glukosa → ADP + glukosa-6-fosfát); tuto reakci katalyzují fosfotransferasy zvané kinasy. Fosforylace se významně podílí na regulaci biologických dějů (viz enzymy – regulace aktivity, též proteinkinasy). Opačný děj (defosforylace), tj. odstranění fosfátové skupiny z organické molekuly, je zprostředkován skupinou enzymů zvaných fosfatasy. Velmi významné jsou fosforylace, jimiž se syntetizuje ATP z ADP (viz fosforylace membránová, fosforylace oxidační a fosforylace substrátová, též fotofosforylace). | |
fosforylace membránováangl. membrane phosphorylation, reakce, při níž je k molekule
ADP připojován (anorganický) fosfátový zbytek: ADP + Pi → ATP + H2O, přičemž tato endergonická reakce získává energii tím, že je spřažena s pasivním transportem protonů biologickou membránou; probíhá tedy na úkor proton-motivní síly. Proces je katalyzován ATP-synthasou. Proton-motivní síla nutná pro tento děj je generována oxidačně-redukčními reakcemi buď v dýchacím řetězci (viz fosforylace oxidační), nebo ve světlé fázi fotosyntézy (viz fotofosforylace). | |
fosforylace oxidačnítéž oxidativní, angl. oxidative phosphorylation, membránová fosforylace u organismů využívajících dýchací řetězec pro generování proton-motivní síly, jeden z nejdůležitějších procesů bioenergetiky. Konečným akceptorem elektronů v dýchacím řetězci může být molekulový kyslík (proto též fosforylace aerobní) nebo jiný extracelulární akceptor elektronů, např. dusičnanový nebo síranový ion (viz anaerobní respirace). Primárním "dodavatelem" elektronů do tohoto procesu jsou u organotrofních organismů organické látky (resp. redukované kofaktory), u chemolithotrofních organismů látky anorganické (sulfan, amoniak, Fe2+ apod.). | |
fosforylace substrátováangl. substrate-level phosphorylation, vedle
membránové fosforylace jediný způsob syntézy nukleosidtrifosfátů, zejména ATP. Jedná se buď o přenos fosfátového zbytku z makroergického substrátu (fosfoenolpyruvátu, 1,3-bisfosfoglycerátu, kreatinfosfátu) na ADP pomocí enzymů z třídy transferas, nebo o obrácenoureakci katalyzovanou ligasou (např. sukcinyl-CoA + GDP + Pi → sukcinát + CoA + GTP, sukcinyl-CoA-ligasa, EC 6.2.1.4, reakce citrátového cyklu). | |
fosforylasyangl. phosphorylases, skupina enzymů z třídy
transferas (EC 2.4.1.-), které katalyzují vznik fosforečných esterů sacharidů přenosem cukerného zbytku z oligosacharidu nebo polysacharidu na anorganický fosfát. Nejdůležitější reakcí tohoto typu je přenos zbytku glukosy z glykogenu na fosfátový ion za vzniku glukosa-1-fosfátu: (glukan)n + Pi → (glukan)n-1 + glukosa-1-fosfát (glykogenfosforylasa, EC 2.4.1.1) | |