Biochemický slovník
Biochemický slovník

Procházet slovníkem pomocí tohoto rejstříku
Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
C |
|---|
cyklus citrátovýtéž Krebsův cyklus, cyklus trikarboxylových kyselin nebo cyklus 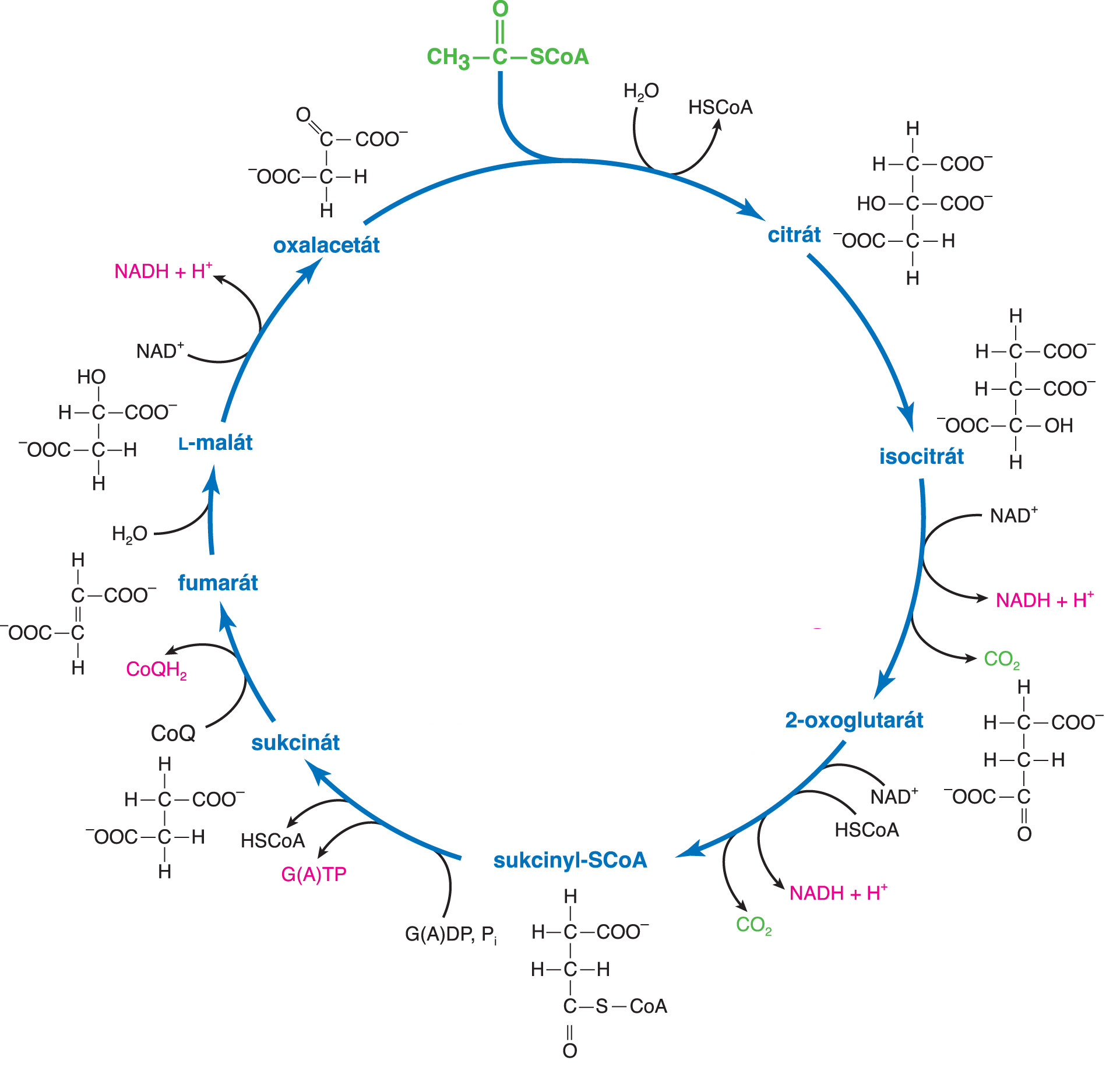 citronové kyseliny, angl. Krebs cycle, hlavní citronové kyseliny, angl. Krebs cycle, hlavní
křižovatkaanabolických a katabolických cest aerobního metabolismu. Jeho sumární rovnice ukazuje, že vstupním metabolitem je acetyl-CoA, který je za součinnosti molekul vody přeměňován na oxid uhličitý; atomy vodíku (v rovnici značené [H], aktivní vodík) jsou navázány na koenzymy oxidoreduktas (NAD+ a CoQ). Následná oxidace NADH a CoQH2 v dýchacím řetězci umožňuje zisk 11 molekul ATP; další makroergická molekula (ATP nebo GTP) vzniká v citrátovém cyklu při rozkladu sukcinyl-CoA reakcí, katalyzovanou ligasou. Význam citrátového cyklu je především energetický. Neslouží však pouze ke katabolické degradaci acetylového zbytku; některé jeho meziprodukty (viz tabulka) jsou prekursory důležitých látek – proto řadíme tento cyklus mezi amfibolické dráhy. U eukaryot je lokalizován v mitochondriích, u prokaryot v cytosolu. Za objev cyklu citronové kyseliny obdržel roku 1953 H. A. Krebs Nobelovu cenu.
| ||||||||||||||||||
cyklus Corihoangl. Cori cycle, komplexní proces, při němž je
laktát, vznikající glykolysou (především v erytrocytech 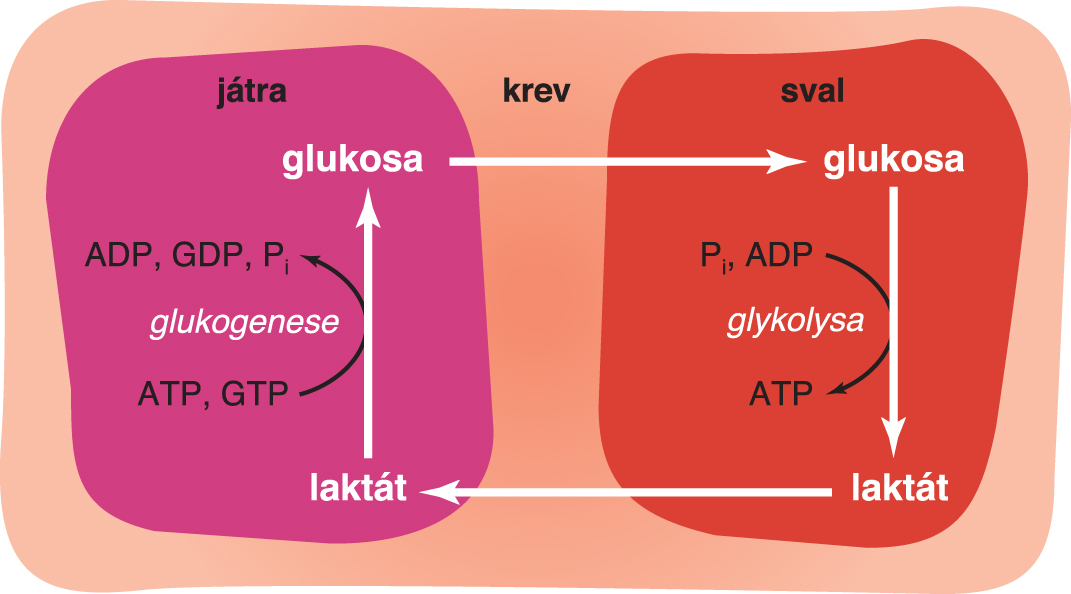 nebo za anaerobních podmínek ve svalech), transportován krví do jater, kde je z něho procesem glukogenese syntetizována glukosa. Ta je opět krví dopravována do tkání, které trvale (erythrocyty) nebo dočasně (svaly) nemohou využívat dýchací řetězec. V játrech je část laktátu aerobně odbourána, čímž se získá ATP, potřebný pro glukogenesi. nebo za anaerobních podmínek ve svalech), transportován krví do jater, kde je z něho procesem glukogenese syntetizována glukosa. Ta je opět krví dopravována do tkání, které trvale (erythrocyty) nebo dočasně (svaly) nemohou využívat dýchací řetězec. V játrech je část laktátu aerobně odbourána, čímž se získá ATP, potřebný pro glukogenesi. | |
cyklus C4-rostlin(psáno C4-rostlin) též Hatchův–Slackův–Kortschackův cyklus [hečúv, slekúv, kortčakúv], 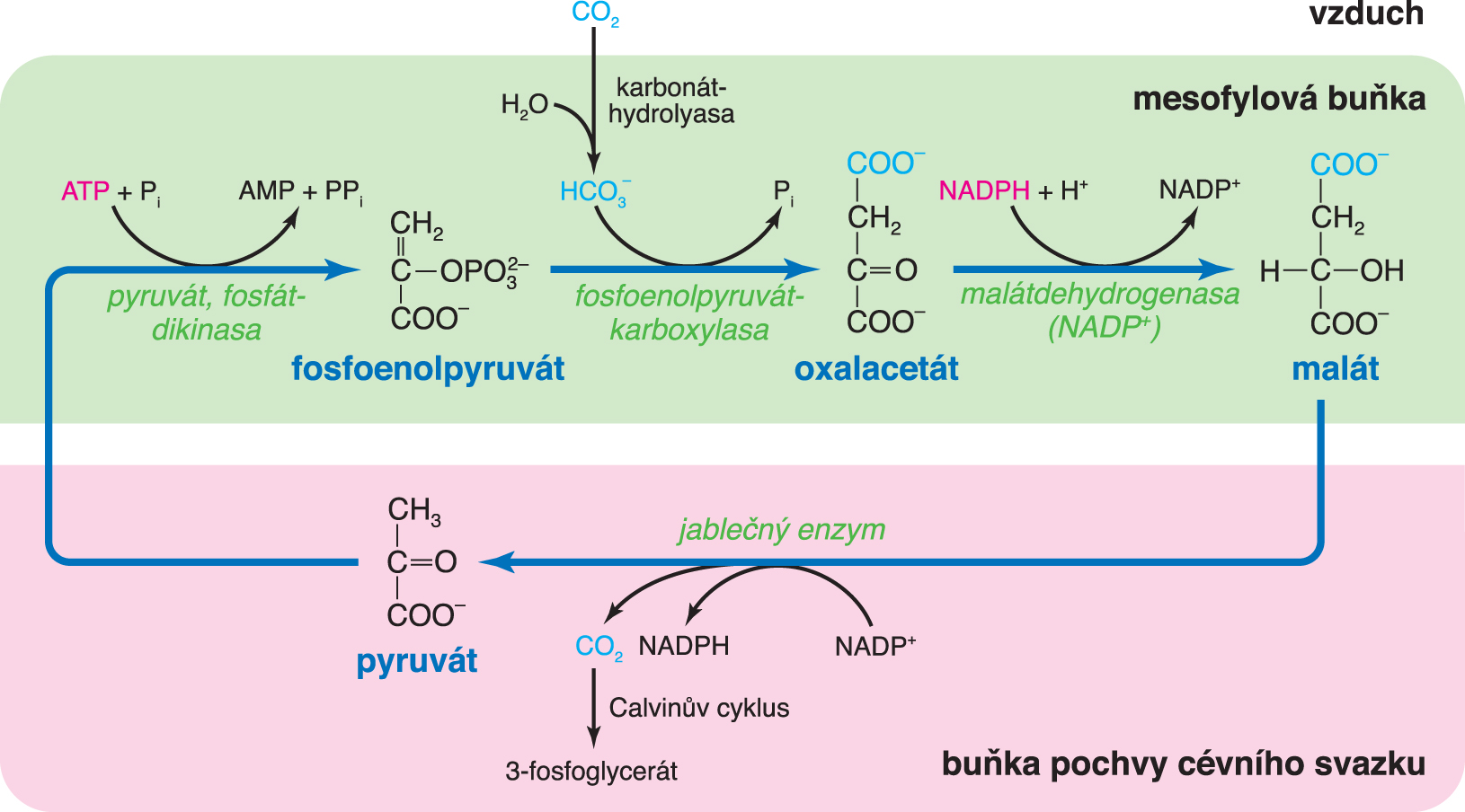 angl. C4-acid cycle, cyklus reakcí při fotosyntetické asimilaci oxidu uhličitého u
C4-rostlin a u CAM-rostlin. Cyklus zahrnuje navázání CO2 na akceptor (fosfoenolpyruvát) a vznik sloučenin se čtyřmi atomy uhlíku (oxalacetát, proto C4), sloužících k transportu navázaného CO2 do buněk, které nejsou v přímém kontaktu s ovzduším (u C4-rostlin), nebo k jeho uložení do zásoby ve vakuolách (malát u CAM-rostlin). Z těchto látek se pak řízeně uvolňuje CO2, který je využíván reakcemi Calvinova cyklu; vznikající pyruvát slouží k regeneraci primárního akceptoru (fosfoenolpyruvátu), čímž se cyklus uzavírá. Řízeným uvolňováním CO2 se, ve srovnání s C3-rostlinami, zvyšuje jeho koncentrace (přesněji parciální tlak) v buňkách, kde probíhá temná fáze fotosyntézy, což zlepšuje jeho využití, neboť snižuje podíl fotorespirace. Rostliny s tímto, z hlediska hospodaření s CO2 výhodným, typem metabolismu (např. kukuřice, cukrová třtina, ananasovník) vyžadují vysokou ozářenost, neboť v temné fázi spotřebovávají více ATP než C3-rostliny. angl. C4-acid cycle, cyklus reakcí při fotosyntetické asimilaci oxidu uhličitého u
C4-rostlin a u CAM-rostlin. Cyklus zahrnuje navázání CO2 na akceptor (fosfoenolpyruvát) a vznik sloučenin se čtyřmi atomy uhlíku (oxalacetát, proto C4), sloužících k transportu navázaného CO2 do buněk, které nejsou v přímém kontaktu s ovzduším (u C4-rostlin), nebo k jeho uložení do zásoby ve vakuolách (malát u CAM-rostlin). Z těchto látek se pak řízeně uvolňuje CO2, který je využíván reakcemi Calvinova cyklu; vznikající pyruvát slouží k regeneraci primárního akceptoru (fosfoenolpyruvátu), čímž se cyklus uzavírá. Řízeným uvolňováním CO2 se, ve srovnání s C3-rostlinami, zvyšuje jeho koncentrace (přesněji parciální tlak) v buňkách, kde probíhá temná fáze fotosyntézy, což zlepšuje jeho využití, neboť snižuje podíl fotorespirace. Rostliny s tímto, z hlediska hospodaření s CO2 výhodným, typem metabolismu (např. kukuřice, cukrová třtina, ananasovník) vyžadují vysokou ozářenost, neboť v temné fázi spotřebovávají více ATP než C3-rostliny. | |
cyklus glyoxylátovýangl. glyoxylate cycle, anaplerotická dráha, sloužící k syntéze sacharidů 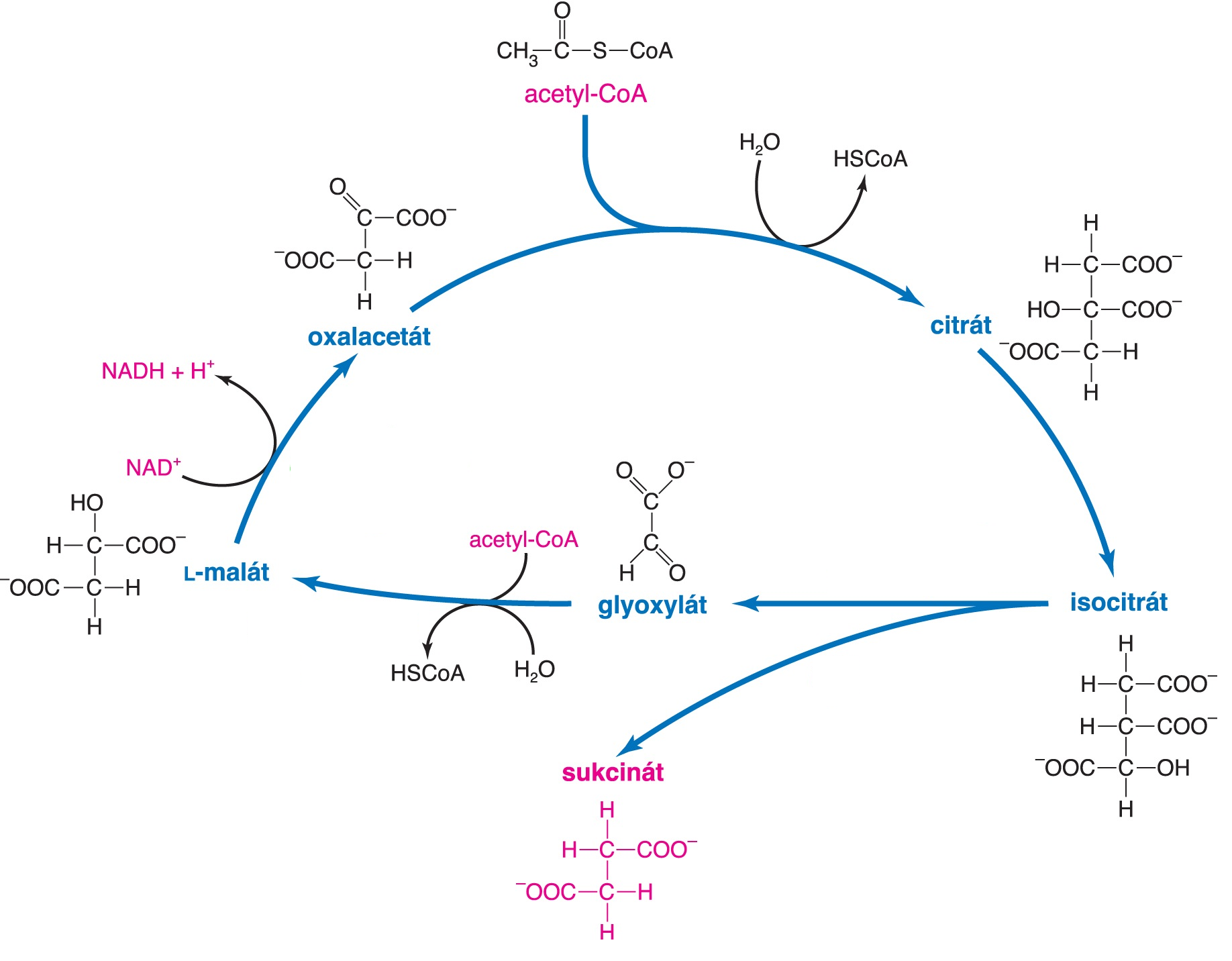 z dvouuhlíkatých sloučenin (z aktivovaného acetátu): z dvouuhlíkatých sloučenin (z aktivovaného acetátu):
Jde o komplexní kondenzační děj, který využívá některých enzymových systémů citrátového cyklu. Klíčovou reakcí je štěpení isocitrátu za vzniku sukcinátu a glyoxylátu; ten v dalším kroku kondenzuje s aktivovaným acetylem. U rostlin a některých mikroorganismů slouží k syntéze oxalacetátu (prekursoru biosyntézy sacharidů procesem glukogenese) z dvouuhlíkatých štěpů (acetyl-CoA), získaných nejčastěji β-oxidací mastných kyselin. Protože živočichové tento cyklus nemají, nemohou z mastných kyselin, uvolněných z lipidů, syntetizovat sacharidy. | |
cyklus metabolickýangl. metabolic cycle, viz metabolické dráhy. | |
cyklus močovinovýtéž ureogenetický, ornithinový nebo Krebsův–Henseleitův cyklus, angl. urea cycle.  Slouží u placentárních savců k eliminaci vysoce toxického
amoniaku, který vzniká především odbouráváním aminokyselin (produkty katabolismu bílkovin). Zdrojem jednoho atomu dusíku v molekule močoviny je zde volný amoniak, zdrojem druhého je aminokyselina L-aspartát (asparagová kyselina): Slouží u placentárních savců k eliminaci vysoce toxického
amoniaku, který vzniká především odbouráváním aminokyselin (produkty katabolismu bílkovin). Zdrojem jednoho atomu dusíku v molekule močoviny je zde volný amoniak, zdrojem druhého je aminokyselina L-aspartát (asparagová kyselina):
Pro přehlednost zde do sumární rovnice nejsou zahrnuty tři molekuly ATP, které se v průběhu tohoto cyklu štěpí a dodávají energii potřebnou k jeho realizaci; dvě molekuly se štěpí na ADP a jedna na AMP, k syntéze jedné molekuly močoviny jsou tedy zapotřebí čtyři jednotky energie ATP. U savců probíhá močovinový cyklus především v játrech (některé reakce v mitochondriích, ostatní v cytosolu buněk). | |
cyklus pentosovýtéž pentosafosfátová cesta, fosfoglukonátová cesta 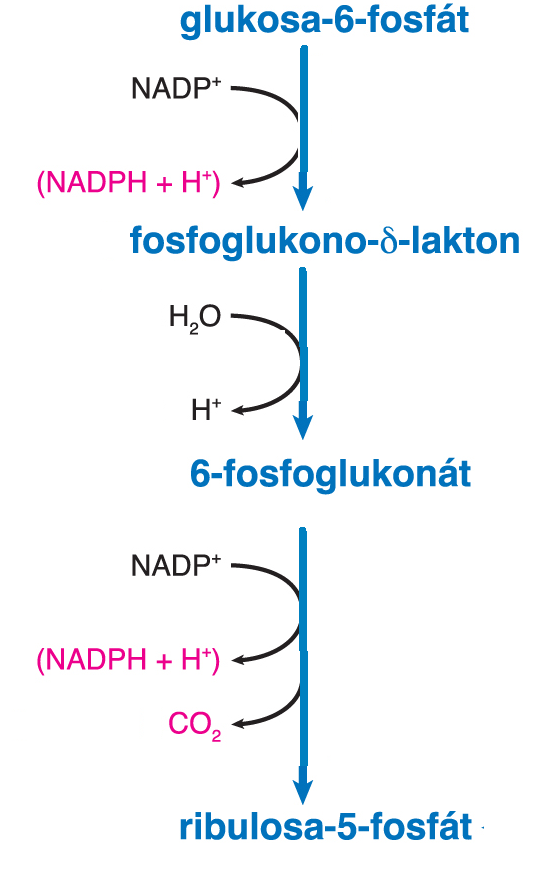 nebo hexosamonofosfátový zkrat, angl. pentose cycle nebo pentose phosphate pathway, komplexní děj, jehož hlavním účelem je získat pro buňku redukční ekvivalenty v podobě
NADPH. V první fázi, zvané oxidační, je výchozí metabolit, glukosa-6-fosfát, dvěma dehydrogenačními kroky převeden na ribulosa-5-fosfát (viz obr.): nebo hexosamonofosfátový zkrat, angl. pentose cycle nebo pentose phosphate pathway, komplexní děj, jehož hlavním účelem je získat pro buňku redukční ekvivalenty v podobě
NADPH. V první fázi, zvané oxidační, je výchozí metabolit, glukosa-6-fosfát, dvěma dehydrogenačními kroky převeden na ribulosa-5-fosfát (viz obr.):
Vzniklá pentosa může být využita pro syntézu nukleotidů, většina je však složitým procesem převedena opět na hexosu; v celkové bilanci této fáze zvané regenerační vzniká ze šesti molekul pentosafosfátu pět molekul hexosafosfátu. Sumární rovnici celého děje (oxidační i regenerační fáze) lze zapsat: | |
cyklus pyruvát-malátovýangl. pyruvate-malate (nebo pyruvate/malate) cycle, složitý proces, pomocí něhož je z pyruvátu 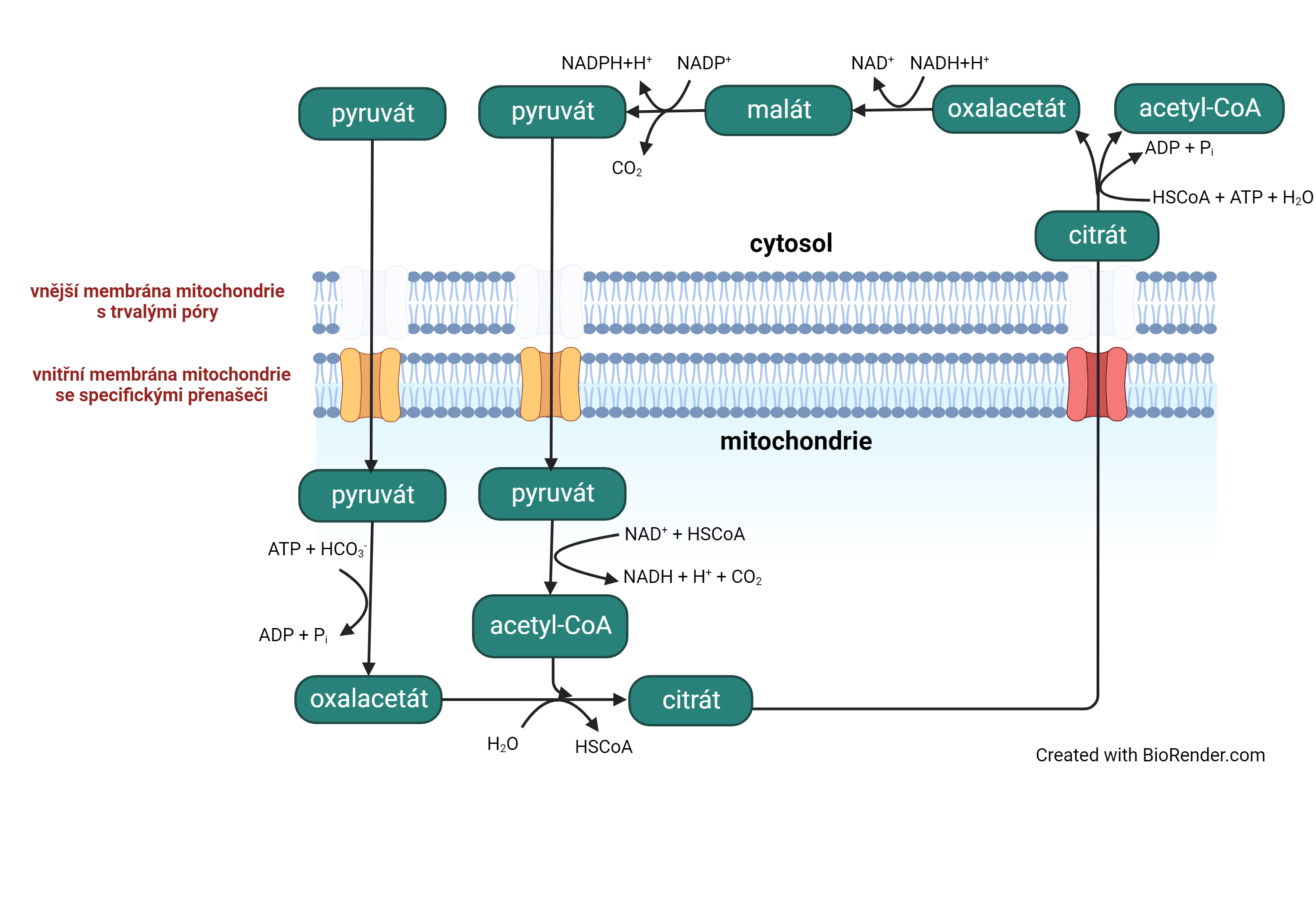 v cytosolu syntetizován acetyl-CoA a současně se získává NADPH. Část děje probíhá v mitochondrii. Sumární rovnici, kde c značí cytosol a m mitochodrii, lze zapsat: v cytosolu syntetizován acetyl-CoA a současně se získává NADPH. Část děje probíhá v mitochondrii. Sumární rovnici, kde c značí cytosol a m mitochodrii, lze zapsat:Cyklus je využíván zejména v situaci, kdy glykolysou vzniká v cytosolu velké množství pyruvátu a NADH a je z nich zapotřebí získat acetyl-CoA a NADPH pro biosyntézu mastných kyselin. V průběhu cyklu se spotřebovávají 2 jednotky ATP, které jsou však nahrazeny reoxidací NADHm v dýchacím řetězci. | |
cyklus zbytečnýtéž psuedocyklus, angl. futile cycle, sekvence reakcí, které mohou vytvářet
metabolický cyklus, obvykle však 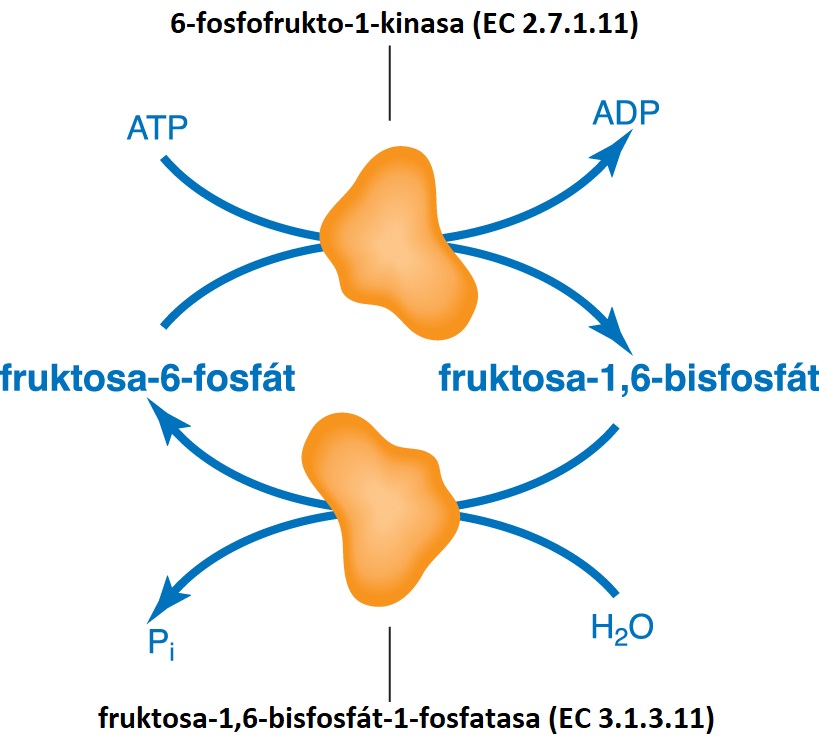 cyklicky neprobíhají, protože celkový zisk užitečné práce by neodpovídal spotřebované energii. Typickým příkladem takového děje je fosforylace fruktosa-6-fosfátu, známá z glykolysy, a hydrolytické štěpení vznikajícího fruktosa-1,6-bisfosfátu, probíhajícího ve stejném kompartmentu (cytosolu) při glukogenesi. Sumárním dějem je tedy štěpení ATP. Za běžných okolností obě reakce neprobíhají současně, neboť jeden z enzymů je vždy inhibován. Některé buňky však využívají tohoto cyklu na produkci tepla. cyklicky neprobíhají, protože celkový zisk užitečné práce by neodpovídal spotřebované energii. Typickým příkladem takového děje je fosforylace fruktosa-6-fosfátu, známá z glykolysy, a hydrolytické štěpení vznikajícího fruktosa-1,6-bisfosfátu, probíhajícího ve stejném kompartmentu (cytosolu) při glukogenesi. Sumárním dějem je tedy štěpení ATP. Za běžných okolností obě reakce neprobíhají současně, neboť jeden z enzymů je vždy inhibován. Některé buňky však využívají tohoto cyklu na produkci tepla. | |
cysteinCys nebo C, angl. cysteine, proteinogenní glukogenní aminokyselina. Může být syntetizován ze serinu, 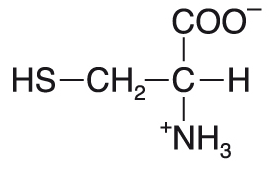 přičemž dodavatelem atomu síry je esenciální aminokyselina methionin; Cys je tedy podmíněně esenciální. Obsahuje thiolovou skupinu (-SH), která je velmi reaktivní a snadno se oxiduje za vzniku disulfidové vazby. Skupina SH dodává postrannímu řetězci Cys polární charakter; disociuje již ve slabě alkalickém prostředí (pKa≈ 8). Cys je součástí tripeptidu glutathionu, který se podílí na řadě intracelulárních oxidačně-redukčních reakcí. Dekarboxylací Cys vzniká významný biogenní amin cysteamin (viz koenzym A). Skupina SH cysteinu je součástí aktivního místa některých proteolytických enzymů (označují se cysteinové neboli SH-proteasy, EC 3.4.22.-; EC 3.4.18.-) přičemž dodavatelem atomu síry je esenciální aminokyselina methionin; Cys je tedy podmíněně esenciální. Obsahuje thiolovou skupinu (-SH), která je velmi reaktivní a snadno se oxiduje za vzniku disulfidové vazby. Skupina SH dodává postrannímu řetězci Cys polární charakter; disociuje již ve slabě alkalickém prostředí (pKa≈ 8). Cys je součástí tripeptidu glutathionu, který se podílí na řadě intracelulárních oxidačně-redukčních reakcí. Dekarboxylací Cys vzniká významný biogenní amin cysteamin (viz koenzym A). Skupina SH cysteinu je součástí aktivního místa některých proteolytických enzymů (označují se cysteinové neboli SH-proteasy, EC 3.4.22.-; EC 3.4.18.-) | |