Biochemický slovník
Biochemický slovník

Speciální | A | Á | B | C | Č | D | Ď | E | É | Ě | F | G | H | CH | I | Í | J | K | L | M | N | Ň | O | Ó | P | Q | R | Ř | S | Š | T | Ť | U | Ú | Ů | V | W | X | Y | Ý | Z | Ž | VŠE
T |
|---|
Tanaka Koichi
| |

technologie fermentačnítéž kvasné technologie, angl. fermentation technologies, souhrnné označení biotechnologických postupů využívajících metabolického aparátu mikroorganismů k výrobě různých produktů, např. alkoholických nápojů, ethanolu, organických kyselin, aminokyselin, vitaminů, steroidních hormonů, antibiotik, ale i droždí nebo krmné biomasy. | |
technologie hybridomovéangl. hybridoma technologies, metodiky využívající
hybridomů, a to nejčastěji k výrobě monoklonálních protilátek. | |
technologie rekombinantníangl. recombinant technologies, soubor metodik využívajících
rekombinantní DNA, a to zejména:
| |
telomerasyangl. telomarases, neobvyklé RNA-dependentní DNA-polymerasy (EC 2.7.7.49) nesoucí si vlastní molekulu RNA jako templát pro tvorbu DNA. Jejich úkolem v buňkách je obnovovat délku zkracujících se telomer. | |
telomeryz řeckého telos = zakončení, angl. telomers, koncové části lineárních chromosomů. Při buněčném dělení se telomery zkracují kvůli neschopnosti DNA-polymeras syntetizovat konce lineárních řetězců DNA. Původní délka se působením enzymu telomerasy může v některých buňkách obnovit (buňky pohlavní, kmenové, rakovinné atd.). Telomery nekódují proteiny a obsahují krátké opakující se sekvence. Postupné zkracování telomer vede ke stárnutí buňky a její programované buněčné smrti, čímž se zabrání nekontrolovatelnému buněčné dělení. Za studium telomer a telomeras získali C. W. Greider, E. H. Blackburn a J. W. Szostak v roce 2009 Nobelovu cenu. | |
templátangl. template, jednořetězcový poly(deoxy)ribonukleotid, využívaný jako zdroj informace při řízené biologické polymeraci (viz řetězec templátový). | |
tenzidyangl. tensides nebo surfactants, látky, které snižují povrchové napětí rozpouštědla (v biochemii vody). Jejich molekuly obsahují polární a nepolární část (viz amfipatické molekuly). V průmyslu i domácnostech se užívají zejména jako čistící a prací prostředky. | |
teorie buněčnáangl. cell theory, teorie formulovaná na počátku 19. století, viz buňka. | |
teorie chemiosmotickáangl. chemiosmotic theory, představa vysvětlující spřažení oxidace redukovaných
kofaktorů (NADH a FADH) se syntézou ATP (ADP + Pi → ATP + H2O) v mitochondriích. Vychází z předpokladu, že energie 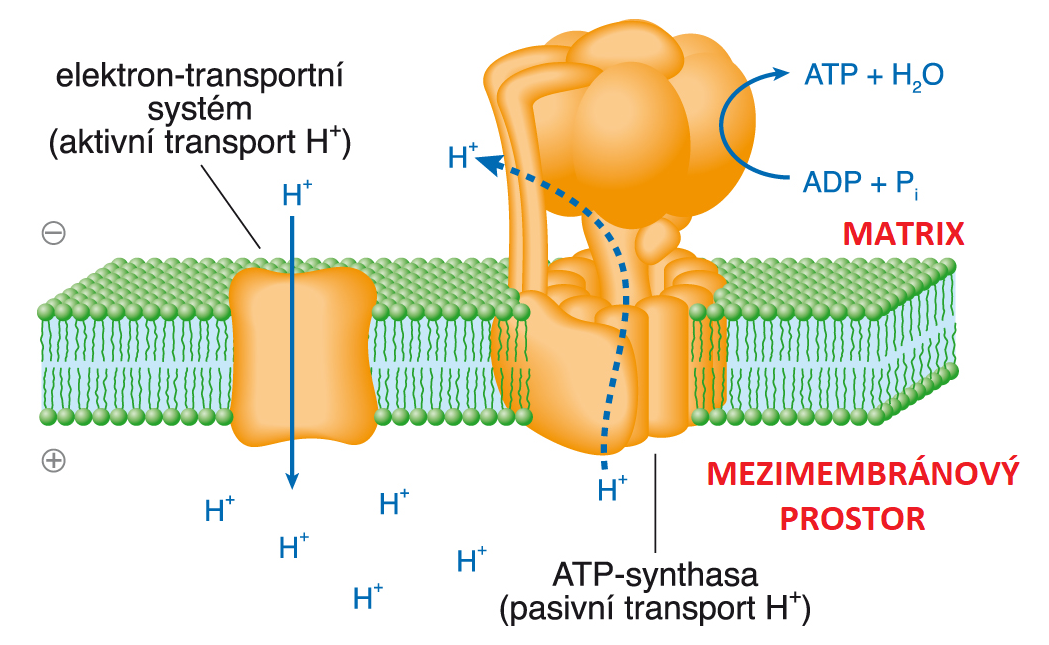 získaná oxidací kofaktorů v dýchacím řetězci (na obr. to je elektron-transportní systém), je nejdříve využita k aktivnímu transportu vodíkových iontů (H+) přes vnitřní membránu mitochondrie do mezimembránového prostoru. Matrix mitochondrie se tak o ionty H+ ochuzuje, a navíc vůči cytosolu získává záporný potenciál. Vzniká specifická forma energie zvaná proton-motivní síla, která tlačíionty H+ zpět do matrix. Zpětný (exergonický, pasivní) transport H+ je umožněn membránovým enzymem ATP-synthasou (EC 7.1.2.2), která uvolněnou energii využívá k výše zmíněné syntéze ATP. Na základě tohoto obecného mechanismu je ATP syntetizován jak v aerobně pracujících mitochondriích, tak na buněčných membránách aerobních a nefermentujících anaerobních mikroorganismů. Proton-motivní síla je generována i při fotosyntéze na membráně thylakoidů, kde je analogicky využívána k syntéze ATP. Za příspěvek k pochopení přenosu biologické energie vypracováním chemiosmotické hypotézy obdržel roku 1978 P. Mitchell Nobelovu cenu. | |
teratogenyangl. teratogens, látky vyvolávající morfologické a funkční odchylky organismu v průběhu vývoje zárodku. | |
terminaceangl. termination, ukončení, pojem užívaný v chemii zejména pro ukončení polymerace nebo řetězové reakce. V biochemii se užívá v souvislosti s ukončením procesu
replikace, transkripce a translace nebo ukončením řetězové reakce vyvolané reaktivními kyslíkovými částicemi. Analogicky se užívají pojmy iniciace a elongace. | |
testosteron
| |
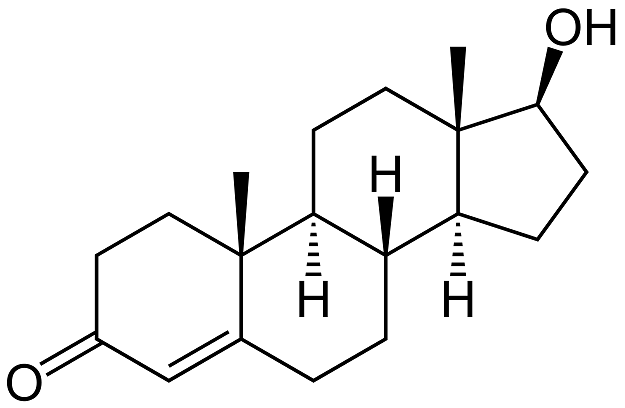 angl. testosterone, mužský pohlavní
angl. testosterone, mužský pohlavní Theorell Axel Hugo Theodor švédský biochemik (1903–1982), získal Nobelovu cenu (za fyziologii a lékařství, 1955) za objevy povahy a mechanismu účinku oxidačních enzymů. Studoval medicínu ve Stockholmu. Po krátkém studiu bakteriologie v Pasteurově ústavu v Paříži pracoval na lékařsko-chemickém ústavu Karolinska Institutet, kde se zabýval vlivem lipidů na sedimentaci krevních elementů. Se studiem oxidoreduktas započal v roce 1933 při svém dvouletém pobytu u O. H. Warburga v Berlíně. Vysvětlil působení flavinových kofaktorů v oxidoreduktasách z droždí. Po návratu na Karolinska Institutet, kde byl v roce 1936 ustanoven přednostou nově vytvořeného biochemického ústavu, se zasloužil o objevy cytochromu-c a řady oxidoreduktas (peroxidasy, katalasy, flavoproteinů a „pyridinových“ proteinů, zvláště alkoholdehydrogenasy). Jeho koníčkem byla hudba; byl členem Švédské královské hudební akademie a ředitelem Stockholmského symfonického orchestru. švédský biochemik (1903–1982), získal Nobelovu cenu (za fyziologii a lékařství, 1955) za objevy povahy a mechanismu účinku oxidačních enzymů. Studoval medicínu ve Stockholmu. Po krátkém studiu bakteriologie v Pasteurově ústavu v Paříži pracoval na lékařsko-chemickém ústavu Karolinska Institutet, kde se zabýval vlivem lipidů na sedimentaci krevních elementů. Se studiem oxidoreduktas započal v roce 1933 při svém dvouletém pobytu u O. H. Warburga v Berlíně. Vysvětlil působení flavinových kofaktorů v oxidoreduktasách z droždí. Po návratu na Karolinska Institutet, kde byl v roce 1936 ustanoven přednostou nově vytvořeného biochemického ústavu, se zasloužil o objevy cytochromu-c a řady oxidoreduktas (peroxidasy, katalasy, flavoproteinů a „pyridinových“ proteinů, zvláště alkoholdehydrogenasy). Jeho koníčkem byla hudba; byl členem Švédské královské hudební akademie a ředitelem Stockholmského symfonického orchestru. | |
thiaminangl. thiamine, viz vitamin B1. | |
threoninThr nebo T, angl. threonine, proteinogenní esenciální glukogenní aminokyselina se 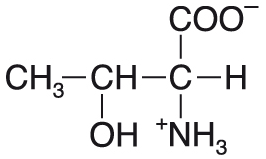 dvěma chirálními centry (stejně jako isoleucin); methylovaný derivát serinu, s nímž má řadu společných funkcí. Ve fosfoproteinech bývá jeho hydroxylová skupina esterifikována zbytkem kyseliny fosforečné. Na postranní řetězec threoninu může být v proteinech O-glykosidovou vazbou navázán oligosacharidový řetězec (O-glykoproteiny). dvěma chirálními centry (stejně jako isoleucin); methylovaný derivát serinu, s nímž má řadu společných funkcí. Ve fosfoproteinech bývá jeho hydroxylová skupina esterifikována zbytkem kyseliny fosforečné. Na postranní řetězec threoninu může být v proteinech O-glykosidovou vazbou navázán oligosacharidový řetězec (O-glykoproteiny). | |
thrombinangl. thrombin, koagulační faktor IIa, enzym, odštěpující na konci koagulační kaskády fibrinopeptidy z fibrinogenu (EC 3.4.21.5). Jeho katalytickou skupinou je serinová proteasa a vykazuje 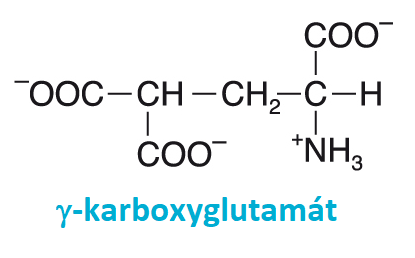 značnou homologii s trávícími enzymy
trypsinem, chymotrypsinem a elastasou. Vzniká rozštěpením molekuly prothrombinu (faktor II) aktivovaným faktorem Xa (EC 3.4.21.6); tato reakce je významně urychlována přítomností aktivovaného faktoru Va a fosfolipidů, je naopak inhibována heparinem. Zatímco prothrombin je tvořen 582 aminokyselinovými zbytky, aktivní thrombin obsahuje pouze 259 zbytků. Syntéza prothrombinu je podobně jako v případě některých dalších koagulačních faktorů závislá na vitaminu K, který se podílí na posttranslační karboxylaci postranních řetězců zbytků glutamové kyseliny (vzniká γ-karboxyglutamylový zbytek). značnou homologii s trávícími enzymy
trypsinem, chymotrypsinem a elastasou. Vzniká rozštěpením molekuly prothrombinu (faktor II) aktivovaným faktorem Xa (EC 3.4.21.6); tato reakce je významně urychlována přítomností aktivovaného faktoru Va a fosfolipidů, je naopak inhibována heparinem. Zatímco prothrombin je tvořen 582 aminokyselinovými zbytky, aktivní thrombin obsahuje pouze 259 zbytků. Syntéza prothrombinu je podobně jako v případě některých dalších koagulačních faktorů závislá na vitaminu K, který se podílí na posttranslační karboxylaci postranních řetězců zbytků glutamové kyseliny (vzniká γ-karboxyglutamylový zbytek). | |
thylakoidyangl. thylakoids, váčky uvnitř
chloroplastů vyšších rostlin a v buňkách sinic; probíhá v nich světlá fáze fotosyntézy. Jsou to malé ploché útvary ohraničené biologickou membránou. Jejich vnitřní prostor se nazývá lumen. V důsledku aktivního transportu vodíkových protonů do thylakoidu je u aktivně fotosyntetizujících chloroplastů pH lumen asi o 3,5 nižší než pH stromatu chloroplastu. V chloroplastech tvoří thylakoidy sloupcovité shluky, zvané grana. | |
thyminangl. thymine, pyrimidinová
báze nukleových kyselin, která je součástí DNA; vzniká methylací 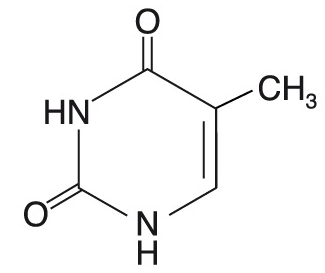 uracilu, který je naopak součástí RNA. Klíčovým bodem biosyntézy thymidinmonofosfátu je methylace deoxyuridinmonofosfátu pomocí kofaktoru N5,N10-methylentetrahydrofolátu; enzym thymidylátsynthasa (EC 2.1.1.45), který tuto reakci katalyzuje, může být inhibován některými cytostatiky (methotrexátem), čímž je zbržděna replikace, a tím i množení nádorových buněk. Pokud jsou v řetězci DNA dva dTMP za sebou, mohou po ozáření rentgenovým nebo ultrafialovým zářením dimerizovat, což vede k mutaci. uracilu, který je naopak součástí RNA. Klíčovým bodem biosyntézy thymidinmonofosfátu je methylace deoxyuridinmonofosfátu pomocí kofaktoru N5,N10-methylentetrahydrofolátu; enzym thymidylátsynthasa (EC 2.1.1.45), který tuto reakci katalyzuje, může být inhibován některými cytostatiky (methotrexátem), čímž je zbržděna replikace, a tím i množení nádorových buněk. Pokud jsou v řetězci DNA dva dTMP za sebou, mohou po ozáření rentgenovým nebo ultrafialovým zářením dimerizovat, což vede k mutaci. | |
thyreotropinthyreoideu-stimulující hormon, TSH, též thyrotropin, angl. thyroid-stimulating hormone, hormon (glykoprotein) uvolňovaný adenohypofýzou, který stimuluje štítnou žlázu, a zvyšuje tak produkci trijodthyroninu (T3) a thyroxinu (T4). Měření hladiny TSH v krvi se využívá k diagnostice onemocnění štítné žlázy. | |
thyroxinangl. thyroxine, zkr. T4, hormon vylučovaný štítnou žlázou. Vzniká posttranslační modifikací bílkoviny thyreoglobulinu, v níž se postupně na zbytky tyrosinu | |
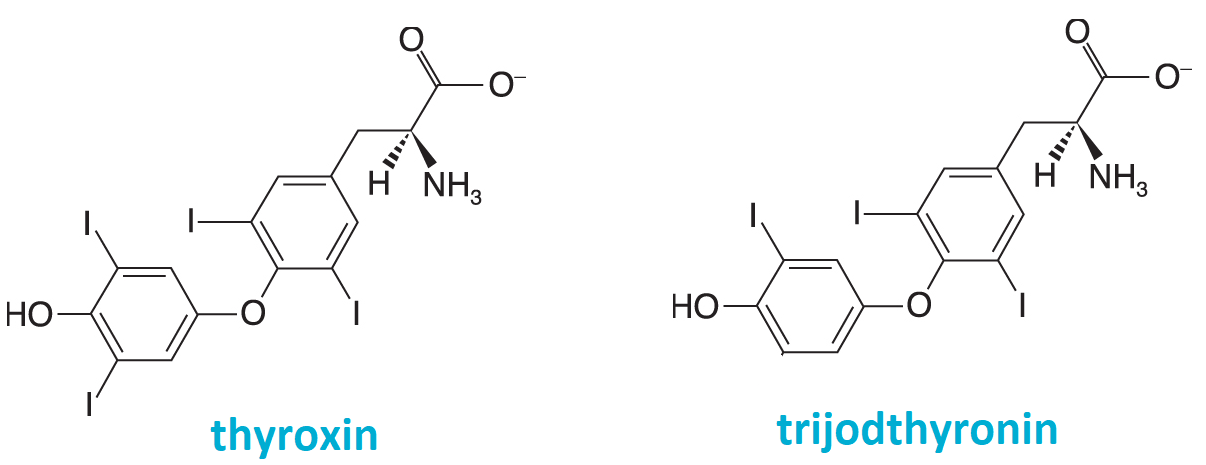 vnášejí čtyři atomy jodu; postranní řetězce
vnášejí čtyři atomy jodu; postranní řetězce Tiselius Arne Wilhelm Kaurin švédský biochemik (1902–1971), Nobelova cena (chemie, 1948) za výzkum elektroforézy a adsorpční analýzy, zejména za objevy týkající se komplexní povahy sérových proteinů. Celý jeho vědecký život je spojen s univerzitou v Upsale, kde jej výrazně ovlivnil T. Svedberg. Již ve své doktorské práci (1930) se zabýval elektroforézou bílkovin. Založil v Upsale Biochemický ústav, který se stal významným centrem studia biochemických separačních metod (elektroforézy, chromatografie atd.). Zastával řadu důležitých funkcí ve vědeckých institucích (předseda IUPAC, předseda Výboru pro udělování Nobelových cen atd.). švédský biochemik (1902–1971), Nobelova cena (chemie, 1948) za výzkum elektroforézy a adsorpční analýzy, zejména za objevy týkající se komplexní povahy sérových proteinů. Celý jeho vědecký život je spojen s univerzitou v Upsale, kde jej výrazně ovlivnil T. Svedberg. Již ve své doktorské práci (1930) se zabýval elektroforézou bílkovin. Založil v Upsale Biochemický ústav, který se stal významným centrem studia biochemických separačních metod (elektroforézy, chromatografie atd.). Zastával řadu důležitých funkcí ve vědeckých institucích (předseda IUPAC, předseda Výboru pro udělování Nobelových cen atd.). | |
tkáňangl. tissue, v živočišných organismech soubor morfologicky a funkčně shodných buněk uspořádaných do pravidelné formace. Rozlišujeme tkáň epitelovou, pojivovou, svalovou a nervovou. Tkáně jednoho nebo více typů vytvářejí
orgány. | |
tlak osmotickýangl. osmotic pressure, tlak vyvolaný tokem rozpouštědla 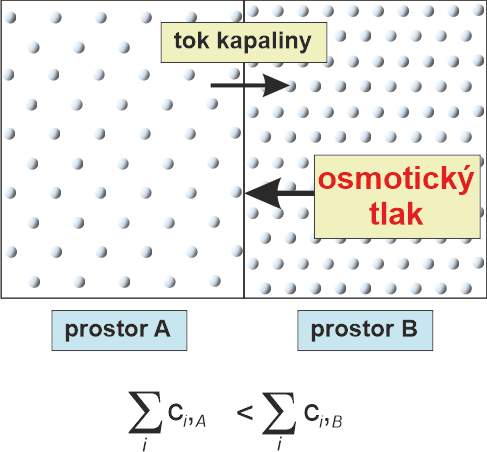 polopropustnou membránou do toho prostoru, v němž je roztok s vyšší koncentrací rozpuštěných molekul nebo iontů. Je dán van´t Hoffovou rovnicí: π = RT Σci, kde π je osmotický tlak, R plynová konstanta, T termodynamická (absolutní) teplota a ci koncentrace jednotlivých osmoticky aktivních částic. Fyziologicky se tato vlastnost vodných roztoků uplatňuje především při filtraci a resorpci vody v ledvinách a při transportu roztoků v rostlinách. Při zacházení zejména s křehkými živočišnými buňkami je třeba dbát na to, aby roztoky, v nichž jsou přechovávány, měly fyziologický osmotický tlak (viz izotonicita).
Za objevy zákonů chemické dynamiky a osmotického tlaku kapalin získal roku 1901 J. H. van't Hoff Nobelovu cenu. polopropustnou membránou do toho prostoru, v němž je roztok s vyšší koncentrací rozpuštěných molekul nebo iontů. Je dán van´t Hoffovou rovnicí: π = RT Σci, kde π je osmotický tlak, R plynová konstanta, T termodynamická (absolutní) teplota a ci koncentrace jednotlivých osmoticky aktivních částic. Fyziologicky se tato vlastnost vodných roztoků uplatňuje především při filtraci a resorpci vody v ledvinách a při transportu roztoků v rostlinách. Při zacházení zejména s křehkými živočišnými buňkami je třeba dbát na to, aby roztoky, v nichž jsou přechovávány, měly fyziologický osmotický tlak (viz izotonicita).
Za objevy zákonů chemické dynamiky a osmotického tlaku kapalin získal roku 1901 J. H. van't Hoff Nobelovu cenu.
| |
Todd Alexandr Robertus britský biochemik (1907–1997), Nobelova cena (chemie, 1957) za práce týkající se nukleotidů a nukleotidových koenzymů. Narodil se v Glasgow (Skotsko) v rodině obchodníka. Vystudoval chemii, zaměřil se na bioorganickou chemii. Syntetizoval řadu biochemicky významných látek (ATP, FAD, UTP), zabýval se strukturou vitaminů B1, B12 a E, jakož i kanabinoidy z hašiše a marihuany. Působil v řadě významných vědeckých funkcí (byl předseda IUPAC, president Royal Society, president poradního sboru Britské vlády pro vědu atd.) V roce 1954 byl povýšen do šlechtického stavu, v roce 1962 jmenován baronem (baron Todd of Trumpington). britský biochemik (1907–1997), Nobelova cena (chemie, 1957) za práce týkající se nukleotidů a nukleotidových koenzymů. Narodil se v Glasgow (Skotsko) v rodině obchodníka. Vystudoval chemii, zaměřil se na bioorganickou chemii. Syntetizoval řadu biochemicky významných látek (ATP, FAD, UTP), zabýval se strukturou vitaminů B1, B12 a E, jakož i kanabinoidy z hašiše a marihuany. Působil v řadě významných vědeckých funkcí (byl předseda IUPAC, president Royal Society, president poradního sboru Britské vlády pro vědu atd.) V roce 1954 byl povýšen do šlechtického stavu, v roce 1962 jmenován baronem (baron Todd of Trumpington). | |
tokoferolyangl. tocopherols, viz vitamin E. | |
transaminaceangl. transamination, reverzibilní přenos aminoskupiny z
2-aminokyseliny (AK) na 2-oxokyselinu (2-OK): AK1 + 2-OK2 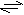 2-OK1 + AK2. Nejčastějším akceptorem aminoskupiny v těchto reakcích je 2-oxoglutarát (meziprodukt citrátového cyklu), z něhož po přijetí aminoskupiny vzniká L-glutamová kyselina. Přenos aminoskupin za vzniku L-glutamátu je častým počátečním krokem odbourávání aminokyselin. Transaminace katalyzují aminotransferasy (starším názvem transaminasy). 2-OK1 + AK2. Nejčastějším akceptorem aminoskupiny v těchto reakcích je 2-oxoglutarát (meziprodukt citrátového cyklu), z něhož po přijetí aminoskupiny vzniká L-glutamová kyselina. Přenos aminoskupin za vzniku L-glutamátu je častým počátečním krokem odbourávání aminokyselin. Transaminace katalyzují aminotransferasy (starším názvem transaminasy). | |
transdukce hormonálního signáluangl. hormone signal transduction, sled událostí popisujících přenos signálu a příslušnou odpověď buněk. Zahrnuje zejména tyto kroky:
| |
transelektronasyangl. transelectronases, nesystémové označení enzymů přenášejících elektrony z donoru na akceptor. Typickými představiteli této skupiny enzymů jsou cytochrom-c-oxidasa (kotvený komplex IV dýchacího řetězce, EC 7.1.1.9) či superoxiddismutasa (EC 1.15.1.1). | |
transferasyangl. transferases, třída enzymů (EC 2.-.-.-), katalyzujících přenos skupiny atomů (SK) z jedné molekuly (donor) na druhou (akceptor): donor-SK + akceptor 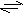 donor + akceptor-SK. Systémový název se tvoří podle schématu donor:akceptor-skupinatransferasa. Transferasy katalyzují řadu velmi významných biochemických reakcí, např. přenos fosfátové skupiny (kinasy), syntézu nukleových kyselin (DNA-polymerasa, RNA-polymerasa) a bílkovin (peptidyltransferasa), štěpení 3-oxoacyl-CoA (thiolasa, thiolytické štěpení), přenos aminoskupin (aminotransferasy). donor + akceptor-SK. Systémový název se tvoří podle schématu donor:akceptor-skupinatransferasa. Transferasy katalyzují řadu velmi významných biochemických reakcí, např. přenos fosfátové skupiny (kinasy), syntézu nukleových kyselin (DNA-polymerasa, RNA-polymerasa) a bílkovin (peptidyltransferasa), štěpení 3-oxoacyl-CoA (thiolasa, thiolytické štěpení), přenos aminoskupin (aminotransferasy). | |
transferrinangl. transferrin, krevní
glykoprotein, jehož funkcí je vázat Fe3+, vytvářet jeho pohotovostní zásobu v krvi a transportovat jej mechanismem endocytózy do buněk. | |
transhydrogenasyangl. transhydrogenases, nesystémové označení
oxidoreduktas přenášejících vodíkové atomy, častější označení dehydrogenasy (viz dehydrogenace). | |
transkripcepřepis, angl. transcription, biosyntéza řetězce
RNA podle templátového řetězce DNA, přičemž jednotlivé nukleotidy jsou připojovány na základě komplementarity (viz báze nukleových kyselin). Klíčovým enzymem této syntézy je RNA-polymerasa. Transkripce (ostatně jako každá polymerační reakce) probíhá ve třech krocích:
| |
transkripce reverzníangl. reverse transcription, proces katalyzovaný enzymem
RNA-dependentní-DNA-polymerasou (EC 2.7.7.49, viz DNA-polymerasa, srov. RNA-polymerasa pro normálnítranskripci), při němž se podle RNA (templát) syntetizuje dvouřetězcová DNA. Jde o významný krok životního cyklu retrovirů, které mají svou genetickou informaci uloženou v molekule RNA, reverzní transkripcí ji transformují do DNA a tu pak inkorporují do chromosomu hostitelské buňky. Reverzní transkripce se využívá v genovém inženýrství, neboť pomocí ní lze funkční mRNA "přepsat" do podoby cDNA (komplementární DNA, genu pro určitou bílkovinu bez intronů), kterou lze po namnožení polymerasovou řetězovou reakcí použít pro přípravu rekombinantní bílkoviny. Do skupiny reverzních transferas řadíme i telomerasu. | |
translaceangl. translation, překlad
genetické informace z řeči nukleotidů do řeči aminokyselin; 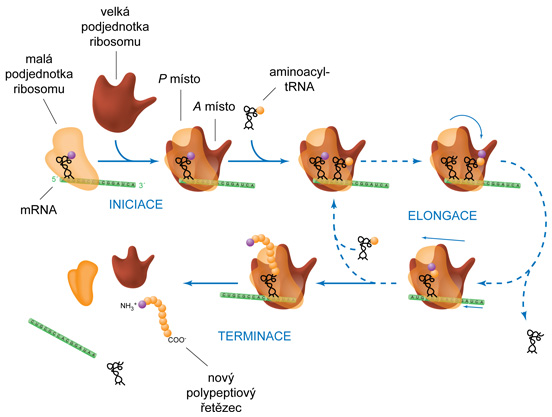 biosyntéza peptidového řetězce podle informace obsažené v mRNA (součást komplexnějšího procesu proteosyntézy). Probíhá na ribosomu za spoluúčasti dalších bílkovinných faktorů. Na velké podjednotce ribosomu jsou dvě vazebná místa: A, do něhož se váže aminoacyl-tRNA, a P, kam se váže peptidyl-tRNA (tRNA, na níž je prostřednictvím C-koncové aminokyseliny navázán peptid, jehož syntéza právě probíhá); obě tRNA jsou prostřednictvím antikodonů navázány na mRNA, která kóduje syntézu daného polypeptidu. Elongační fáze translace probíhá v pěti krocích: biosyntéza peptidového řetězce podle informace obsažené v mRNA (součást komplexnějšího procesu proteosyntézy). Probíhá na ribosomu za spoluúčasti dalších bílkovinných faktorů. Na velké podjednotce ribosomu jsou dvě vazebná místa: A, do něhož se váže aminoacyl-tRNA, a P, kam se váže peptidyl-tRNA (tRNA, na níž je prostřednictvím C-koncové aminokyseliny navázán peptid, jehož syntéza právě probíhá); obě tRNA jsou prostřednictvím antikodonů navázány na mRNA, která kóduje syntézu daného polypeptidu. Elongační fáze translace probíhá v pěti krocích:
Proces translace je iniciován vazbou Met-tRNAMet do místa P ribosomu a prostřednictvím interakce kodon-antikodon k iniciačnímu kodonu mRNA; do místa A se pak naváže další aminoacyl-tRNA (viz výše popsané kroky). K ukončení translace (terminaci) dochází v případě, že do místa A se dostane STOP-kodon. | |
translokace skupinováangl. group translocation, typ transportu látek biologickou membránou, při němž dochází k chemické modifikaci přenášené částice. Typickým příkladem je transport acylových zbytků z cytosolu do mitochondrie pomocí karnitinu. | |
translokasyangl. translocases, nová enzymová třída zavedená v srpnu 2018 (EC 7.-.-.-). Jsou sem zařazeny enzymy, zajišťující aktivní transport částic biologickými membránami. Třída se dělí na podtřídy podle toho, jaká
částice se transportuje, zda proton (podtřída 7.1), anorganické kationty (7.2),
anorganické anionty (7.3), aminokyseliny a peptidy (7.4), sacharidy (7.5) nebo
ostatní částice (7.6). O zařazení do
skupin rozhoduje typ exergonické chemické reakce, která aktivnímu transportu
dodává energii; ve skupině 7.X.1 jsou to oxidačně-redukční reakce (např.
transport protonů kotvenými komplexy, EC 7.1.1.2, EC 7.1.1.8 a EC 7.1.1.9),
ve skupině 7.X.2 je dodavatelem energie hydrolýza nukleosidtrifosfátu,
nejčastěji ATP (např. ATP-synthasa EC 7.1.2.2 nebo Na,K-ATPasa EC 7.2.2.13) atd.
Doposud (září 2022) však bohužel nebyly zveřejněny závazné systémové názvy
enzymů této třídy a nadále se používají systémové názvy odpovídající „starému“ zařazení enzymů ve třídách 1 až 6. Pozn.: Před rokem 2018 byl pojem translokasy používán pro integrální membránové bílkoviny katalyzující antiport (někdy též symport) částic membránou. K nejznámějším patřila ATP-ADP-translokasa, která zajišťuje výměnu ATP za ADP mezi matrix mitochondrie a cytosolem. | |
transport aktivníangl. active trransport, proces, při němž je částice (molekula nebo ion) přenášena biologickou membránou proti gradientu svého elektrochemického potenciálu, tedy tak, že její energie roste. Energie vzrůstá proto, že v kompartmentu, kam se částice transportuje, má vyšší koncentraci, 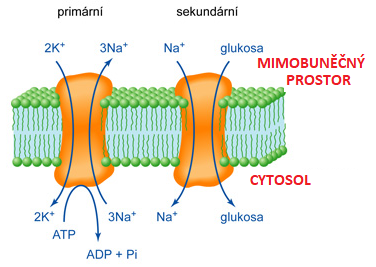 nebo proto, že transportovaný ion tam má v důsledku membránového potenciálu vyšší elektrostatickou energii. Aktivní transport je vždy zajišťován integrálními membránovými bílkovinami. Patří k základním projevům života, neboť zajišťuje stabilitu složení vnitřního prostředí buněk, přičemž toto složení je trvale odlišné od složení extracelulárního prostředí. nebo proto, že transportovaný ion tam má v důsledku membránového potenciálu vyšší elektrostatickou energii. Aktivní transport je vždy zajišťován integrálními membránovými bílkovinami. Patří k základním projevům života, neboť zajišťuje stabilitu složení vnitřního prostředí buněk, přičemž toto složení je trvale odlišné od složení extracelulárního prostředí.Energie, potřebná pro tento transport, se získává:
| |
transport bílkovin kotranslační a posttranslačnítéž targeting (target, čti target = cíl), angl. cotranslational and posttranslational transport of proteins, cílení nebo směrování bílkovin, souhrnný název procesů umožňujících správnou a cílenou lokalizaci bílkovin. Ke kotranslačnímu transportu dochází při translaci na hrubém
endoplasmatickém retikulu, kdy hydrofobní vedoucí sekvence umožňuje, aby bílkovina prošla membránou do endoplasmatického retikula a tam byla dále upravována (zejména odštěpením vedoucí sekvence a glykosylací). Posttranslační transport bílkovin (vylučování extracelulárních bílkovin z buňky, přesuny mezi jednotlivými organelami atd.) je zajišťován různými váčky a následně mechanismy exocytózy. | |
transport membránovýangl. membrane transport, v biochemii soubor procesů umožňujících látkám různého typu překonat barieru
biologické membrány. Z hlediska mechanismu přenosu částic rozlišujeme transport
| |
transport pasivníangl. passive transport, transport částic (molekul nebo iontů)
biologickou membránou ve směru, kdy elektrochemický potenciál částice klesá; nevyžaduje tedy dodávání energie (srov. aktivní transport). Pasivní transport může být realizován všemi postupy popsanými u hesla transport membránový. | |
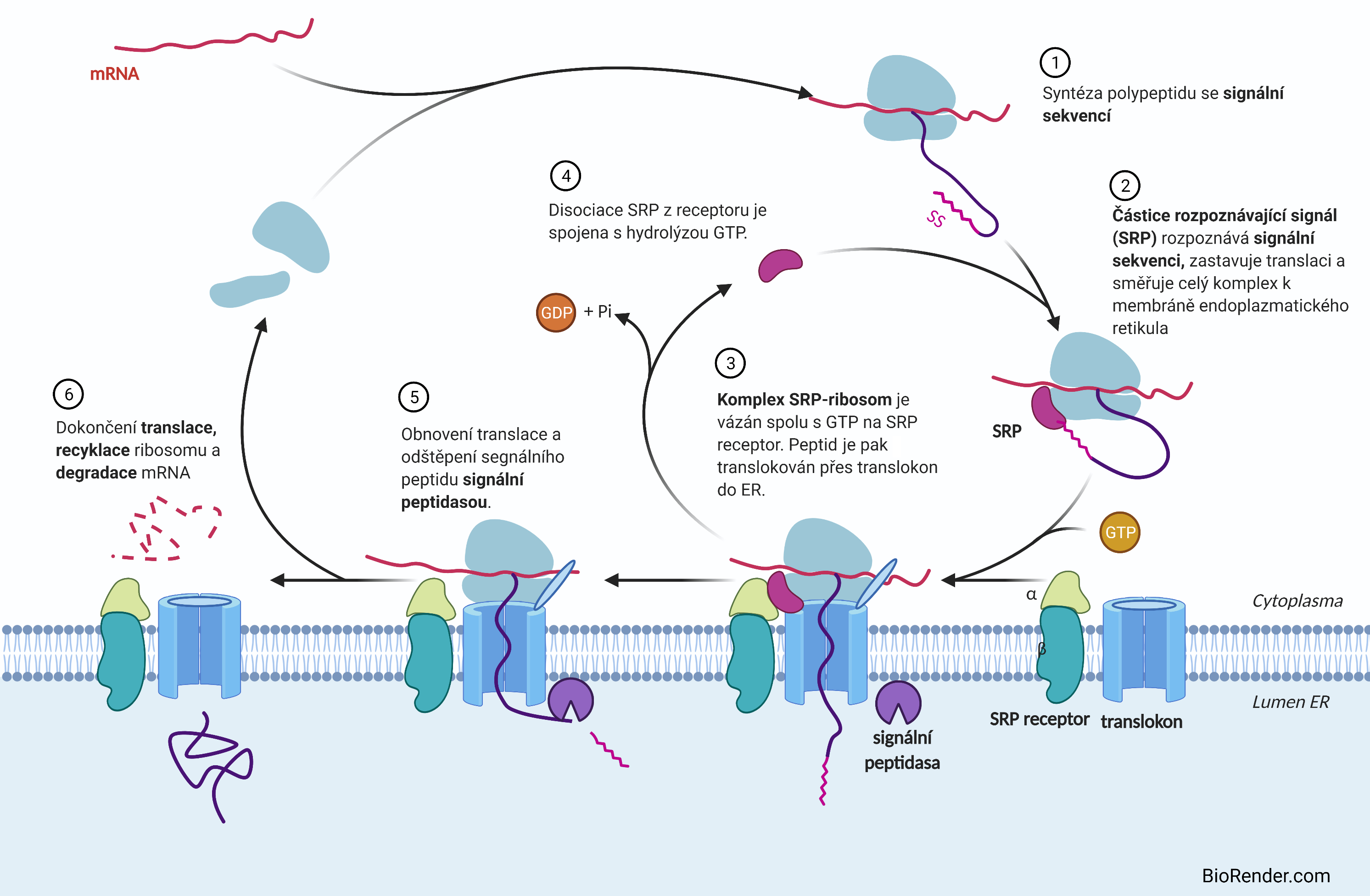
transport proteinů do endoplasmatického retikulaangl. transport of proteins into the endoplasmic reticulum, proces umožňující polypeptidovému řetězci vnikajícímu pomocí ribosomu na vnějším povrchu hrubého endoplasmatického retikula (ER) proniknout do jeho lumen, kde pak podléhá dalším modifikacím. Pomocí hrubého ER jsou syntetizovány prakticky všechny transmembránové proteiny a probíhají v něm také mnohé posttranslační modifikace (např. N-glykosylace). Všechny tyto proteiny mají na N-konci polypeptidu signální sekvenci, na kterou se naváže na částice rozpoznávající signál (SRP proteiny) a umožní jí překonat membránovou barieru.
| |
tráveníangl. digestion, ve fyziologii příjem potravy, její rozklad (digesce) a vstřebávání (resorpce) vznikajících nízkomolekulárních látek v trávicím traktu živočichů; v biochemii hydrolytický enzymový rozklad komplexních, často nerozpustných látek (živin) v trávicím traktu živočichů nebo v lysosomech (buněčné trávení). | |
triacylglycerolyangl. triacylglycerols, zkr. TAG, estery
glycerolu a vyšších mastných kyselin (monoacylglyceroly a diacylglyceroly se vyskytují pouze jako meziprodukty metabolismu triacylglycerolů nebo fosfatidátů). U živočichů jsou v nich obsaženy převážně nevětvené mastné kyseliny se sudým počtem atomů uhlíku. Směsi triacylglycerolů se nazývají tuky (někdy též neutrální tuky, též lipidy v užším slova smyslu). TAG patří k základním živinám. V tenkém střevu jsou částečně hydrolyzovány, především na monoacylglyceroly, které jsou pak v buňkách střevního epitelu reesterifikovány a po vytvoření komplexů se specifickými proteiny, cholesterolem a fosfolipidy vylučovány do krevního řečiště v podobě lipoproteinových částic. TAG obsažené v buňkách tukové tkáně (adipocytech) jsou u živočichů významnou zásobárnou energie, jejich syntéza (lipogeneze) a odbourávání jsou řízeny hormonálně. Mnoho rostlin (řepka, slunečnice, len, olivy aj.) vytváří v semenech zásoby TAG ve formě rostlinných olejů. | |
triglyceridyangl. triglycerids, zastaralé, leč v medicině stále ještě užívané označení
triacylglycerolů. | |
trijodthyroninangl. triiodothironine, zkr. T3, viz thyroxin. | |
tRNAtransferová RNA, angl. transfer RNA, jednořetězcová RNA, obvykle tvořená 70 až 85 zbytky ribonukleotidů 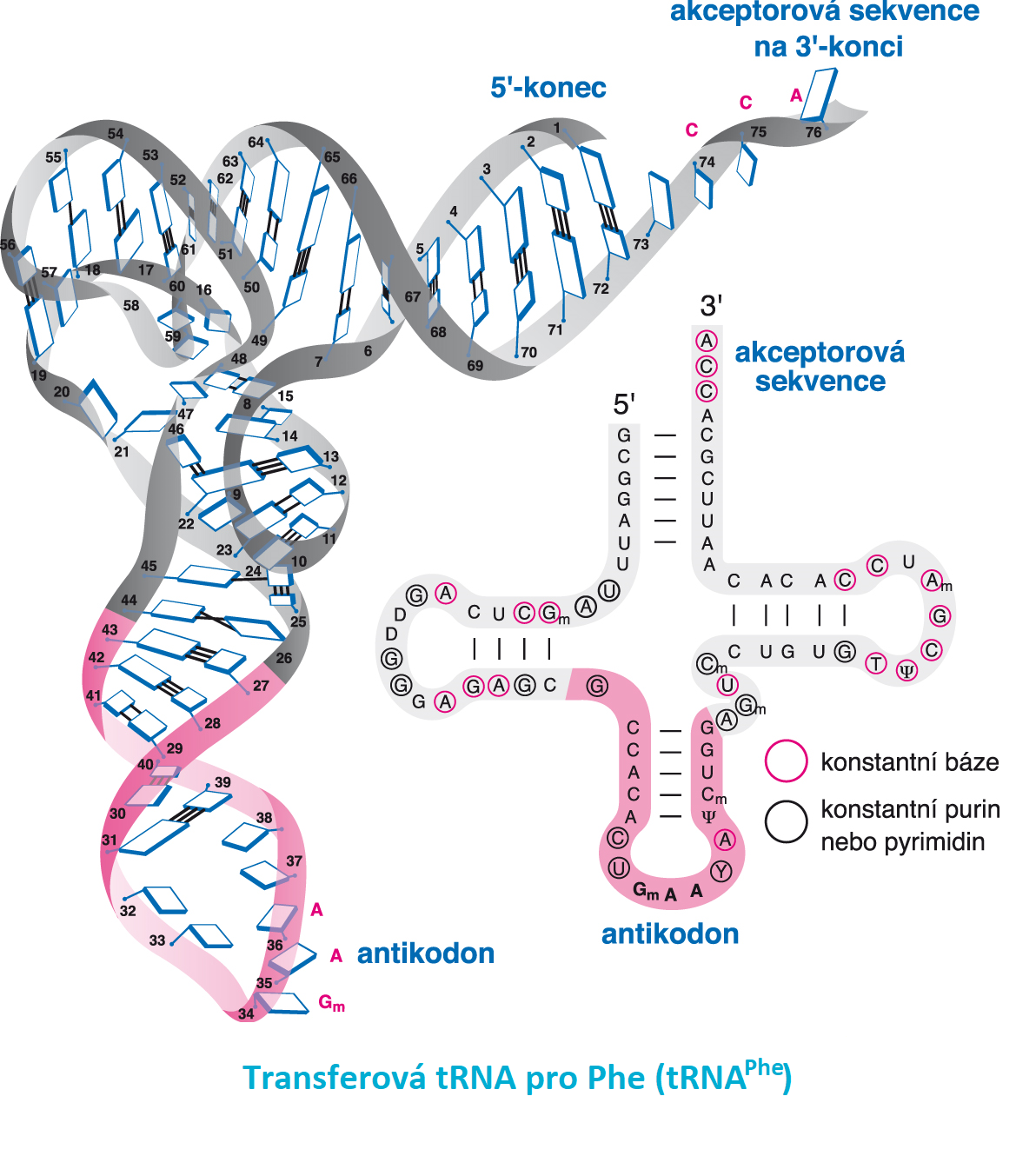 (molekulová hmotnost přibližně 2,4 · 104 Da). Obsahuje relativně mnoho minoritních bází jako důsledek posttranskripčních modifikací. Má poměrně přesně definovanou prostorovou strukturu (sekundární struktura podobná jetelovému listu, terciární struktura připomínající písmeno L). Obvykle je v buňce 50 až 70 molekulových druhů tRNA; důvodem této heterogenity je (molekulová hmotnost přibližně 2,4 · 104 Da). Obsahuje relativně mnoho minoritních bází jako důsledek posttranskripčních modifikací. Má poměrně přesně definovanou prostorovou strukturu (sekundární struktura podobná jetelovému listu, terciární struktura připomínající písmeno L). Obvykle je v buňce 50 až 70 molekulových druhů tRNA; důvodem této heterogenity je
Funkčně nejdůležitějšími částmi tRNA jsou antikodon (trojice nukleotidů komplementární k určitému kodonu a určující specifitu tRNA pro danou aminokyselinu) a vždy stejná trojice nukleotidů CCA na 3′-konci, kam se na volnou OH-skupinu koncové ribosy esterovou vazbou váže aminokyselina za vzniku aminoacyl-tRNA (viz aminoacyl-tRNA-ligasy). | |
trofikaangl. trophics, výživa, (z řeckého trofé = výživa), též způsoby výživy organismů, resp. způsoby přijímání uhlíku a získávání energie (viz organismy – rozdělení podle způsobu výživy). | |
tropinyangl. tropins, skupina
hormonů vylučovaná adenohypofýzou a řídící činnost dalších žláz a buněk. Tak thyreotropin řídí činnost štítné žlázy, gonadotropiny činnost pohlavních žláz, adrenokortikotropin činnost kůry nadledvin atd. | |
tropomyosinangl. tropomyosine, bílkovina nekovalentně spojená s aktinem a tvořící s ním základ svalových filament a mikrofilament cytoskeletu ostatních typů eukaryotních buněk. | |
trypsinangl. trypsin, EC 3.4.21.4, serinová
proteasa syntetizovaná ve formě proenzymu trypsinogenu ve slinivce břišní všech obratlovců. Trypsinogen je vylučován do dvanáctníku, kde je aktivován odštěpením N-koncového hexapeptidu. Specificky štěpí peptidové vazby vycházející z karboxylové skupiny bazických aminokyselin argininu a lysinu. Jeho optimum pH je v mírně alkalické oblasti odpovídající hodnotě pH dvanáctníku. | |
tryptofanTrp nebo W, angl. tryptophan, aromatická esenciální ketogenní proteinogenní 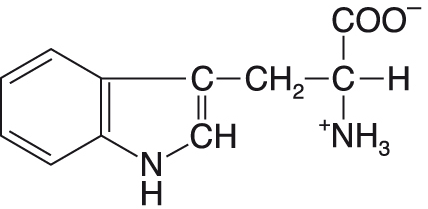 aminokyselina. Jeho objemný postranní řetězec je významnou strukturní součástí bílkovin. Má význačné fluorescenční vlastnosti, čehož se využívá při studiu prostorového uspořádání okolí jeho indolové skupiny. Ačkoliv je v bílkovinách poměrně málo zastoupen, je velmi významný z hlediska výživy. Je prekursorem několika důležitých látek: nikotinamidu (viz niacin) a biogenních aminů tryptaminu a serotoninu. aminokyselina. Jeho objemný postranní řetězec je významnou strukturní součástí bílkovin. Má význačné fluorescenční vlastnosti, čehož se využívá při studiu prostorového uspořádání okolí jeho indolové skupiny. Ačkoliv je v bílkovinách poměrně málo zastoupen, je velmi významný z hlediska výživy. Je prekursorem několika důležitých látek: nikotinamidu (viz niacin) a biogenních aminů tryptaminu a serotoninu. | |
Tsien Roger Yonchien
| |

tubulin | |
tukyangl. lipids, triviální označení
triacylglycerolů, v technologii se někdy rozlišují tuky (za pokojové teploty pevné, s vyšším obsahem nasycených mastných kyselin) a oleje (kapalné). | |
tyrosinTyr nebo Y, angl. tyrosine, aromatická proteinogenní ketogenní aminokyselina, 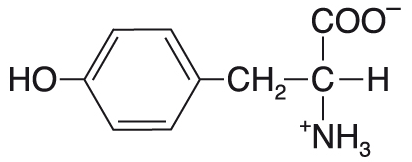 podmíněně esenciální (vzniká hydroxylací esenciální aminokyseliny fenylalaninu). Je prekursorem řady významných látek především hormonální povahy: katecholaminů (dopaminu, noradrenalinu a adrenalinu) a hormonů štítné žlázy (trijodtyroninu a thyroxinu); jeho dekarboxylačním produktem je biogenní amin tyramin. Jeho fenolová hydroxylová skupina může být fosforylována působením protein-tyrosin kinas (EC 2.7.10.-); tato modifikace je významným prostředkem regulace aktivity řady enzymů, ukazuje se, že mnoho onkogenů kóduje právě tento typ kinas (viz tyrosinkinasové receptory). Fenolová skupina OH má vlastnosti slabé kyseliny; při pH vyšším než přibližně 10 disociuje za vzniku fenolátového iontu. podmíněně esenciální (vzniká hydroxylací esenciální aminokyseliny fenylalaninu). Je prekursorem řady významných látek především hormonální povahy: katecholaminů (dopaminu, noradrenalinu a adrenalinu) a hormonů štítné žlázy (trijodtyroninu a thyroxinu); jeho dekarboxylačním produktem je biogenní amin tyramin. Jeho fenolová hydroxylová skupina může být fosforylována působením protein-tyrosin kinas (EC 2.7.10.-); tato modifikace je významným prostředkem regulace aktivity řady enzymů, ukazuje se, že mnoho onkogenů kóduje právě tento typ kinas (viz tyrosinkinasové receptory). Fenolová skupina OH má vlastnosti slabé kyseliny; při pH vyšším než přibližně 10 disociuje za vzniku fenolátového iontu. | |